| MTR | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Identifiers | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Aliases | MTR, HMAG, MS, cblG, 5-methyltetrahydrofolate-homocysteine methyltransferase | ||||||||||||||||||||||||||||||||||||||||||||||||||
| External IDs | OMIM: 156570 MGI: 894292 HomoloGene: 37280 GeneCards: MTR | ||||||||||||||||||||||||||||||||||||||||||||||||||
| EC number | 2.1.1.13 | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Wikidata | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||




Methionine synthase (MS, MeSe, MTR) is responsible for the regeneration of methionine from homocysteine. In humans it is encoded by the MTR gene (5-methyltetrahydrofolate-homocysteine methyltransferase).[5][6] Methionine synthase forms part of the S-adenosylmethionine (SAMe) biosynthesis and regeneration cycle,[7] and is the enzyme responsible for linking the cycle to one-carbon metabolism via the folate cycle. There are two primary forms of this enzyme, the Vitamin B12 (cobalamin)-dependent (MetH) and independent (MetE) forms,[8] although minimal core methionine synthases that do not fit cleanly into either category have also been described in some anaerobic bacteria.[9] The two dominant forms of the enzymes appear to be evolutionary independent and rely on considerably different chemical mechanisms.[10] Mammals and other higher eukaryotes express only the cobalamin-dependent form. In contrast, the distribution of the two forms in Archaeplastida (plants and algae) is more complex. Plants exclusively possess the cobalamin-independent form,[11] while algae have either one of the two, depending on species.[12] Many different microorganisms express both the cobalamin-dependent and cobalamin-independent forms.[13]
Mechanism

Methionine synthase catalyzes the final step in the regeneration of methionine (Met) from homocysteine (Hcy). Both the cobalamin-dependent and cobalamin-independent forms of the enzyme carry out the same overall chemical reaction, the transfer of a methyl group from 5-methyltetrahydrofolate (N5-MeTHF) to homocysteine, yielding tetrahydrofolate (THF) and methionine.[8] Methionine synthase is the only mammalian enzyme that metabolizes N5-MeTHF to regenerate the active cofactor THF. In the cobalamin-dependent (MetH) form of the enzyme, the reaction proceeds by two steps in a preferred ordered sequential mechanism.[14] The physiological resting state of the enzyme is thought to contain the enzyme-bound(Cob) cofactor in the methylcobalamin form, with the cobalt atom in the formal +3 valence state (Cob(III)-Me). The cobalamin is then demethylated by zinc-activated thiolate homocysteine, generating methionine and reducing the cofactor to a Cob(I) state. When in the Cob(I) form, the enzyme-bound cofactor is now able to abstract a methyl group from activated 5-methyltetrahydrofolate (N5-MeTHF), yielding tetrahydrofolate (THF) and regenerating the methylcoalamin form of the enzyme.[15]

Under physiological conditions, approximately once every 2000 catalytic turnovers the Co(I) may be oxidized into inactive Co(II) in cob-dependent MetH. To account for this effect, the protein contains a self-reactivation mechanism, a reductive methylation process that uses S-adenosylmethionine as a distinct methyl donor. In humans, the enzyme is reduced in this process by methionine synthase reductase (MTRR), which consists of flavodoxin-like and ferrodoxin-NADP+ oxidoreductase (FNR)-like domains.[16] In many bacteria, the reduction is carried out by a single domain flavodoxin protein.[17] The reductase protein is responsible for transfer of an electron from a reduced FMN cofactor to the inactive Cob(II), which enables regeneration of the active methylcobalamin enzyme via methyl transfer from S-adenosylmethionine to the reduced Cob(I) intermediate.[18] This process is known as the reactivation cycle, and is thought to be gated from the normal catalytic cycle by large-scale conformational rearrangements within the enzyme.[19] Because the oxidation of Cob(I) inevitably shuts down cob-dependent methionine synthase activity, defects or deficiencies in methionine synthase reductase have been implicated in some of the disease associations for methionine synthase deficiency.[20]
The mechanism of the cobalamin-independent (MetE) form, by contrast, proceeds through a direct methyl transfer from the activated N5-MeTHF to zinc thiolate homocysteine. Although the mechanism is considerably simpler, the direct transfer reaction is much less favorable than the cobalamin-mediated reactions and as a result the turnover rate for MetE is ~100x slower than that of MetH. As it does not contain the cobalamin cofactor, the cobalamin-independent enzyme is not prone to oxidative inactivation [21][8][22][23]
Structure

High-resolution structures have been solved by X-ray crystallography for intact MetE both in the absence and presence of substrates[23][22] and for fragments of MetH,[24][25][26][27] although no structural description exists of a fully intact MetH enzyme. The available structures and accompanying bioinformatic analysis indicate minimal similarity in the overall structure, although there are similarities within the substrate-binding sites themselves.[28] Cob-dependent MetH is divided into 4 separate domains. The domains, from N- to C-terminus, are denoted homocysteine binding (Hcy domain), N5-methylTHF binding (MTHF domain) Cobalamin-binding (Cob domain) and the S-adenosymethionine-binding or reactivation domain. The reactivation domain binds SAM and is the site of interaction with flavodoxin or Methionine Synthase Reductase during the reactivation cycle of the enzyme.[17][16][20] The cobalamin-binding domain contains two subdomains, with the cofactor bound to the Rossman-fold B12-binding subdomain, which is in turn capped by the other subdomain, the four-helix bundle cap subdomain.[25] The four-helix bundle serves to protect the cobalamin cofactor from unwanted reactivity, but can significantly change conformations to expose the cofactor allow it access to the other substrates during turnover.[26] Both the Hcy and N5-MeTHF domains adopt a TIM barrel architecture; the Hcy domain contains the zinc-binding site, which in MetH consists of three cysteine residues coordinated to a zinc ion which in turn binds and activates Hcy. The N5-MeTHF binding domain binds and activates N5-MeTHF via a hydrogen bonding network with several asparagine, arginine, and aspartic acid residues. During turnover, the enzyme undergoes significant conformational changes that involve moving the Cob-domain back and forth from the Hcy domain to the N5-MeTHF domain in order for the two methyl transfer reactions to proceed.[24]
The cob-independent MetE consists of two TIM-barrel domains that bind homocysteine and N5-MeTHF individually. The two domains adopt a face-to-face double barrel architecture, which requires a "closing" of the structure upon binding of both substrates to enable the direct methyl transfer.[22] Substrate-binding strategies are similar to MetH, although in the case of MetE the zinc atom is instead coordinated to two cysteines, a histidine and a glutamate,[23] for which an example is shown on the right.
Biochemical function
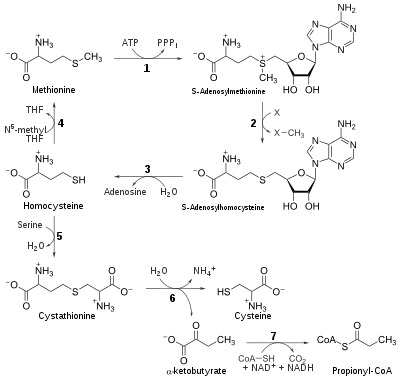
In humans the enzyme's main purpose is to regenerate Met in the S-adenosylmethionine (SAM) cycle. The SAM cycle in a single turnover consumes Met and ATP and generates Hcy, and can involve any of a number of critical enzymatic reactions that use S-adenosylmethionine as the source of an active methyl group for methylation of nucleic acids, histones, phospholipids and various proteins.[29][30] As such, methionine synthase serves an essential function by allowing the SAM cycle to perpetuate without a constant influx of Met. As a secondary effect, methionine synthase also serves to maintain low levels of Hcy and, because methionine synthase is one of the few enzymes that used N5-MeTHF as a substrate, to indirectly maintain THF levels.[31][32]
In bacteria and plants, methionine synthase serves a dual purpose of both perpetuating the SAM cycle and catalyzing the final synthetic step in the de novo synthesis of Met, which is one of the 20 canonical amino acids.[33][11] While the chemical reaction is exactly the same for both processes, the overall function is distinct from methionine synthase in humans because Met is an essential amino acid that is not synthesized de novo in the body.[34]
Clinical significance
Mutations in the MTR gene have been identified as the underlying cause of methylcobalamin deficiency complementation group G, or methylcobalamin deficiency cblG-type.[5] Deficiency or deregulation of the enzyme due to deficient methionine synthase reductase can directly result in elevated levels of homocysteine (hyperhomocysteinemia), which is associated with blindness, neurological symptoms, and birth defects.[35][36] Methionine synthase reductase (MTRR) or methylene-tetrahydrofolate reductase (MTHFR) deficiencies can also result in the condition. Most cases of methionine synthase deficiency are symptomatic within 2 years of birth with many patients rapidly developing severe encephalopathy.[37] One consequence of reduced methionine synthase activity that is measurable by routine clinical blood tests is megaloblastic anemia.
Genetics
Several cblG-associated polymorphisms in the MTR gene have been identified.[38]
- 2756D→G (Asp919Gly)
- 3804C→T (Pro1137Leu)
- Δ2926A-2928T (ΔIle881)
See also
References
- 1 2 3 GRCh38: Ensembl release 89: ENSG00000116984 - Ensembl, May 2017
- 1 2 3 GRCm38: Ensembl release 89: ENSMUSG00000021311 - Ensembl, May 2017
- ↑ "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- ↑ "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- 1 2 "MTR 5-methyltetrahydrofolate-homocysteine methyltransferase (Homo sapiens)". Entrez. 19 May 2009. Retrieved 24 May 2009.
- ↑ Li YN, Gulati S, Baker PJ, Brody LC, Banerjee R, Kruger WD (December 1996). "Cloning, mapping and RNA analysis of the human methionine synthase gene". Human Molecular Genetics. 5 (12): 1851–1858. doi:10.1093/hmg/5.12.1851. PMID 8968735.
- ↑ Banerjee RV, Matthews RG (March 1990). "Cobalamin-dependent methionine synthase". FASEB Journal. 4 (5): 1450–1459. doi:10.1096/fasebj.4.5.2407589. hdl:2027.42/154369. PMID 2407589. S2CID 8210250.
- 1 2 3 Matthews RG, Smith AE, Zhou ZS, Taurog RE, Bandarian V, Evans JC, Ludwig M (2003). "Cobalamin-Dependent and Cobalamin-Independent Methionine Synthases: Are There Two Solutions to the Same Chemical Problem?". Helvetica Chimica Acta. 86 (12): 3939–3954. doi:10.1002/hlca.200390329.
- ↑ Deobald D, Hanna R, Shahryari S, Layer G, Adrian L (February 2020). "Identification and characterization of a bacterial core methionine synthase". Scientific Reports. 10 (1): 2100. Bibcode:2020NatSR..10.2100D. doi:10.1038/s41598-020-58873-z. PMC 7005905. PMID 32034217.
- ↑ Pejchal R, Ludwig ML (February 2005). Stroud RM (ed.). "Cobalamin-independent methionine synthase (MetE): a face-to-face double barrel that evolved by gene duplication". PLOS Biology. 3 (2): e31. doi:10.1371/journal.pbio.0030031. PMC 539065. PMID 15630480.
- 1 2 Ravanel S, Gakière B, Job D, Douce R (June 1998). "The specific features of methionine biosynthesis and metabolism in plants". Proceedings of the National Academy of Sciences of the United States of America. 95 (13): 7805–7812. Bibcode:1998PNAS...95.7805R. doi:10.1073/pnas.95.13.7805. PMC 22764. PMID 9636232.
- ↑ Helliwell KE, Wheeler GL, Leptos KC, Goldstein RE, Smith AG (October 2011). "Insights into the evolution of vitamin B12 auxotrophy from sequenced algal genomes". Molecular Biology and Evolution. 28 (10): 2921–2933. doi:10.1093/molbev/msr124. PMID 21551270.
- ↑ Zydowsky TM (1986). "Stereochemical analysis of the methyl transfer catalyzed by cobalamin-dependent methionine synthase from Escherichia coli B". Journal of the American Chemical Society. 108 (11): 3152–3153. doi:10.1021/ja00271a081.
- ↑ Banerjee RV, Frasca V, Ballou DP, Matthews RG (December 1990). "Participation of cob(I) alamin in the reaction catalyzed by methionine synthase from Escherichia coli: a steady-state and rapid reaction kinetic analysis". Biochemistry. 29 (50): 11101–11109. doi:10.1021/bi00502a013. PMID 2271698.
- ↑ Chemistry and biochemistry of B12. Ruma Banerjee. New York: Wiley. 1999. ISBN 0-471-25390-1. OCLC 40397055.
{{cite book}}: CS1 maint: others (link) - 1 2 Yamada K, Gravel RA, Toraya T, Matthews RG (June 2006). "Human methionine synthase reductase is a molecular chaperone for human methionine synthase". Proceedings of the National Academy of Sciences of the United States of America. 103 (25): 9476–9481. Bibcode:2006PNAS..103.9476Y. doi:10.1073/pnas.0603694103. PMC 1480432. PMID 16769880.
- 1 2 Hall DA, Jordan-Starck TC, Loo RO, Ludwig ML, Matthews RG (September 2000). "Interaction of flavodoxin with cobalamin-dependent methionine synthase". Biochemistry. 39 (35): 10711–10719. doi:10.1021/bi001096c. PMID 10978155.
- ↑ Jarrett JT, Hoover DM, Ludwig ML, Matthews RG (September 1998). "The mechanism of adenosylmethionine-dependent activation of methionine synthase: a rapid kinetic analysis of intermediates in reductive methylation of Cob(II)alamin enzyme". Biochemistry. 37 (36): 12649–12658. doi:10.1021/bi9808565. PMID 9730838.
- ↑ Jarrett JT, Huang S, Matthews RG (April 1998). "Methionine synthase exists in two distinct conformations that differ in reactivity toward methyltetrahydrofolate, adenosylmethionine, and flavodoxin". Biochemistry. 37 (16): 5372–5382. doi:10.1021/bi9730893. PMID 9548919.
- 1 2 Wolthers KR, Scrutton NS (June 2007). "Protein interactions in the human methionine synthase-methionine synthase reductase complex and implications for the mechanism of enzyme reactivation". Biochemistry. 46 (23): 6696–6709. doi:10.1021/bi700339v. PMID 17477549.
- ↑ Zhang Z, Tian C, Zhou S, Wang W, Guo Y, Xia J, et al. (December 2012). "Mechanism-based design, synthesis and biological studies of N⁵-substituted tetrahydrofolate analogs as inhibitors of cobalamin-dependent methionine synthase and potential anticancer agents". European Journal of Medicinal Chemistry. 58: 228–236. doi:10.1016/j.ejmech.2012.09.027. PMID 23124219.
- 1 2 3 Ubhi DK, Robertus JD (February 2015). "The cobalamin-independent methionine synthase enzyme captured in a substrate-induced closed conformation". Journal of Molecular Biology. 427 (4): 901–909. doi:10.1016/j.jmb.2014.12.014. PMID 25545590.
- 1 2 3 Koutmos M, Pejchal R, Bomer TM, Matthews RG, Smith JL, Ludwig ML (March 2008). "Metal active site elasticity linked to activation of homocysteine in methionine synthases". Proceedings of the National Academy of Sciences of the United States of America. 105 (9): 3286–3291. Bibcode:2008PNAS..105.3286K. doi:10.1073/pnas.0709960105. PMC 2265165. PMID 18296644.
- 1 2 Evans JC, Huddler DP, Hilgers MT, Romanchuk G, Matthews RG, Ludwig ML (March 2004). "Structures of the N-terminal modules imply large domain motions during catalysis by methionine synthase". Proceedings of the National Academy of Sciences of the United States of America. 101 (11): 3729–3736. Bibcode:2004PNAS..101.3729E. doi:10.1073/pnas.0308082100. PMC 374312. PMID 14752199.
- 1 2 Drennan CL, Huang S, Drummond JT, Matthews RG, Ludwig ML (December 1994). "How a protein binds B12: A 3.0 A X-ray structure of B12-binding domains of methionine synthase". Science. 266 (5191): 1669–1674. doi:10.1126/science.7992050. PMID 7992050.
- 1 2 Bandarian V, Pattridge KA, Lennon BW, Huddler DP, Matthews RG, Ludwig ML (January 2002). "Domain alternation switches B(12)-dependent methionine synthase to the activation conformation". Nature Structural Biology. 9 (1): 53–56. doi:10.1038/nsb738. PMID 11731805. S2CID 10529695.
- ↑ Datta S, Koutmos M, Pattridge KA, Ludwig ML, Matthews RG (March 2008). "A disulfide-stabilized conformer of methionine synthase reveals an unexpected role for the histidine ligand of the cobalamin cofactor". Proceedings of the National Academy of Sciences of the United States of America. 105 (11): 4115–4120. Bibcode:2008PNAS..105.4115D. doi:10.1073/pnas.0800329105. PMC 2393809. PMID 18332423.
- ↑ Pejchal R, Ludwig ML (February 2005). "Cobalamin-independent methionine synthase (MetE): a face-to-face double barrel that evolved by gene duplication". PLOS Biology. 3 (2): e31. doi:10.1371/journal.pbio.0030031. PMC 539065. PMID 15630480.
- ↑ Broderick JB, Duffus BR, Duschene KS, Shepard EM (April 2014). "Radical S-adenosylmethionine enzymes". Chemical Reviews. 114 (8): 4229–4317. doi:10.1021/cr4004709. PMC 4002137. PMID 24476342.
- ↑ Shane B (June 2008). "Folate and vitamin B12 metabolism: overview and interaction with riboflavin, vitamin B6, and polymorphisms". Food and Nutrition Bulletin. 29 (2 Suppl): S5–S16. doi:10.1177/15648265080292S103. PMID 18709878. S2CID 21493797.
- ↑ Watkins D, Ru M, Hwang HY, Kim CD, Murray A, Philip NS, et al. (July 2002). "Hyperhomocysteinemia due to methionine synthase deficiency, cblG: structure of the MTR gene, genotype diversity, and recognition of a common mutation, P1173L". American Journal of Human Genetics. 71 (1): 143–153. doi:10.1086/341354. PMC 384971. PMID 12068375.
- ↑ Ghergurovich JM, Xu X, Wang JZ, Yang L, Ryseck RP, Wang L, Rabinowitz JD (November 2021). "Methionine synthase supports tumour tetrahydrofolate pools". Nature Metabolism. 3 (11): 1512–1520. doi:10.1038/s42255-021-00465-w. PMC 9284419. PMID 34799699. S2CID 244450216.
- ↑ Ferla MP, Patrick WM (August 2014). "Bacterial methionine biosynthesis". Microbiology. 160 (Pt 8): 1571–1584. doi:10.1099/mic.0.077826-0. PMID 24939187.
- ↑ Hesse H, Hoefgen R (June 2003). "Molecular aspects of methionine biosynthesis". Trends in Plant Science. 8 (6): 259–262. doi:10.1016/S1360-1385(03)00107-9. PMID 12818659.
- ↑ Banerjee RV, Matthews RG (March 1990). "Cobalamin-dependent methionine synthase". FASEB Journal. 4 (5): 1450–1459. doi:10.1096/fasebj.4.5.2407589. hdl:2027.42/154369. PMID 2407589. S2CID 8210250.
- ↑ Kim J, Kim H, Roh H, Kwon Y (April 2018). "Causes of hyperhomocysteinemia and its pathological significance". Archives of Pharmacal Research. 41 (4): 372–383. doi:10.1007/s12272-018-1016-4. PMID 29552692. S2CID 3986295.
- ↑ Outteryck O, de Sèze J, Stojkovic T, Cuisset JM, Dobbelaere D, Delalande S, et al. (July 2012). "Methionine synthase deficiency: a rare cause of adult-onset leukoencephalopathy". Neurology. 79 (4): 386–388. doi:10.1212/WNL.0b013e318260451b. PMID 22786600. S2CID 207121496.
- ↑ Gulati S, Baker P, Li YN, Fowler B, Kruger W, Brody LC, Banerjee R (December 1996). "Defects in human methionine synthase in cblG patients". Human Molecular Genetics. 5 (12): 1859–1865. doi:10.1093/hmg/5.12.1859. PMID 8968736.
Further reading
- Ludwig ML, Matthews RG (1997). "Structure-based perspectives on B12-dependent enzymes". Annual Review of Biochemistry. 66: 269–313. doi:10.1146/annurev.biochem.66.1.269. PMID 9242908.
- Matthews RG, Sheppard C, Goulding C (April 1998). "Methylenetetrahydrofolate reductase and methionine synthase: biochemistry and molecular biology". European Journal of Pediatrics. 157 (Suppl 2): S54–S59. doi:10.1007/PL00014305. hdl:2027.42/42254. PMID 9587027. S2CID 8709190.
- Garovic-Kocic V, Rosenblatt DS (August 1992). "Methionine auxotrophy in inborn errors of cobalamin metabolism". Clinical and Investigative Medicine. 15 (4): 395–400. PMID 1516297.
- O'Connor DL, Moriarty P, Picciano MF (1992). "The impact of iron deficiency on the flux of folates within the mammary gland". International Journal for Vitamin and Nutrition Research. 62 (2): 173–180. PMID 1517041.
- Everman BW, Koblin DD (March 1992). "Aging, chronic administration of ethanol, and acute exposure to nitrous oxide: effects on vitamin B12 and folate status in rats". Mechanisms of Ageing and Development. 62 (3): 229–243. doi:10.1016/0047-6374(92)90109-Q. PMID 1583909. S2CID 11766691.
- Vassiliadis A, Rosenblatt DS, Cooper BA, Bergeron JJ (August 1991). "Lysosomal cobalamin accumulation in fibroblasts from a patient with an inborn error of cobalamin metabolism (cblF complementation group): visualization by electron microscope radioautography". Experimental Cell Research. 195 (2): 295–302. doi:10.1016/0014-4827(91)90376-6. PMID 2070814.
- Li YN, Gulati S, Baker PJ, Brody LC, Banerjee R, Kruger WD (December 1996). "Cloning, mapping and RNA analysis of the human methionine synthase gene". Human Molecular Genetics. 5 (12): 1851–1858. doi:10.1093/hmg/5.12.1851. PMID 8968735.
- Gulati S, Baker P, Li YN, Fowler B, Kruger W, Brody LC, Banerjee R (December 1996). "Defects in human methionine synthase in cblG patients". Human Molecular Genetics. 5 (12): 1859–1865. doi:10.1093/hmg/5.12.1859. PMID 8968736.
- Leclerc D, Campeau E, Goyette P, Adjalla CE, Christensen B, Ross M, et al. (December 1996). "Human methionine synthase: cDNA cloning and identification of mutations in patients of the cblG complementation group of folate/cobalamin disorders". Human Molecular Genetics. 5 (12): 1867–1874. doi:10.1093/hmg/5.12.1867. PMID 8968737.
- Chen LH, Liu ML, Hwang HY, Chen LS, Korenberg J, Shane B (February 1997). "Human methionine synthase. cDNA cloning, gene localization, and expression". The Journal of Biological Chemistry. 272 (6): 3628–3634. doi:10.1074/jbc.272.6.3628. PMID 9013615.
- Wilson A, Leclerc D, Saberi F, Campeau E, Hwang HY, Shane B, et al. (August 1998). "Functionally null mutations in patients with the cblG-variant form of methionine synthase deficiency". American Journal of Human Genetics. 63 (2): 409–414. doi:10.1086/301976. PMC 1377317. PMID 9683607.
- Salomon O, Rosenberg N, Zivelin A, Steinberg DM, Kornbrot N, Dardik R, et al. (2002). "Methionine synthase A2756G and methylenetetrahydrofolate reductase A1298C polymorphisms are not risk factors for idiopathic venous thromboembolism". The Hematology Journal. 2 (1): 38–41. doi:10.1038/sj.thj.6200078. PMID 11920232.
- Watkins D, Ru M, Hwang HY, Kim CD, Murray A, Philip NS, et al. (July 2002). "Hyperhomocysteinemia due to methionine synthase deficiency, cblG: structure of the MTR gene, genotype diversity, and recognition of a common mutation, P1173L". American Journal of Human Genetics. 71 (1): 143–153. doi:10.1086/341354. PMC 384971. PMID 12068375.
- De Marco P, Calevo MG, Moroni A, Arata L, Merello E, Finnell RH, et al. (2002). "Study of MTHFR and MS polymorphisms as risk factors for NTD in the Italian population". Journal of Human Genetics. 47 (6): 319–324. doi:10.1007/s100380200043. PMID 12111380.
- Doolin MT, Barbaux S, McDonnell M, Hoess K, Whitehead AS, Mitchell LE (November 2002). "Maternal genetic effects, exerted by genes involved in homocysteine remethylation, influence the risk of spina bifida". American Journal of Human Genetics. 71 (5): 1222–1226. doi:10.1086/344209. PMC 385102. PMID 12375236.
- Zhu H, Wicker NJ, Shaw GM, Lammer EJ, Hendricks K, Suarez L, et al. (March 2003). "Homocysteine remethylation enzyme polymorphisms and increased risks for neural tube defects". Molecular Genetics and Metabolism. 78 (3): 216–221. doi:10.1016/S1096-7192(03)00008-8. PMID 12649067.
External links
- GeneReviews/NCBI/NIH/UW entry on Disorders of Intracellular Cobalamin Metabolism
- ENZYME: EC 2.1.1.13
- 5-Methyltetrahydrofolate-Homocysteine+S-Methyltransferase at the U.S. National Library of Medicine Medical Subject Headings (MeSH)
PDB gallery | |
|---|---|
|
