| Feline coronavirus | |
|---|---|
| Virus classification | |
| (unranked): | Virus |
| Realm: | Riboviria |
| Kingdom: | Orthornavirae |
| Phylum: | Pisuviricota |
| Class: | Pisoniviricetes |
| Order: | Nidovirales |
| Family: | Coronaviridae |
| Genus: | Alphacoronavirus |
| Subgenus: | Tegacovirus |
| Species: | |
| Virus: | Feline coronavirus |
| Strains[1] | |
| |
Feline coronavirus (FCoV) is a positive-stranded RNA virus that infects cats worldwide.[2] It is a coronavirus of the species Alphacoronavirus 1, which includes canine coronavirus (CCoV) and porcine transmissible gastroenteritis coronavirus (TGEV). FCoV has two different forms: feline enteric coronavirus (FECV), which infects the intestines, and feline infectious peritonitis virus (FIPV), which causes the disease feline infectious peritonitis (FIP).
Feline coronavirus is typically shed in feces by healthy cats, and transmitted by the fecal-oral route to other cats.[3] In environments with multiple cats, the transmission rate is much higher compared to single-cat environments.[2] The virus is insignificant until mutations cause it to be transformed from FECV to FIPV.[2] FIPV causes feline infectious peritonitis, for which treatment is generally symptomatic and palliative only. The drug GS-441524 shows promise as an antiviral treatment for FIP, but at the moment it still requires further research.[4] The drug GC376 is also being studied and developed.
Prevalence
Feline coronavirus is found in cat populations around the world. The only known exceptions are on the Falkland Islands and the Galapagos, where studies found no occurrences of FCoV antibodies in cats tested.[5][6]
Virology

Feline enteric coronavirus (FECV)
Feline enteric coronavirus is responsible for an infection of the mature gastrointestinal epithelial cells[7] (see also enterocytes, brush border, microvilli, villi). This intestinal infection has few outward signs, and is usually chronic. The virus is excreted in the feces of the healthy carrier, and can be detected by polymerase chain reaction (PCR) of feces or by PCR testing of rectal samples.[7]
Cats living in groups can infect each other with different strains of the virus during visits to a communal litter tray. Some cats are resistant to the virus and can avoid infection or even becoming carriers, while others may become FECV carriers.[7]
Feline infectious peritonitis virus (FIPV) and Feline infectious peritonitis
The virus becomes feline infectious peritonitis virus (FIPV) when random errors occur in the virus infecting an enterocyte, causing the virus to mutate from FECV to FIPV.[7]
In their pre-domestication natural state, cats are solitary animals and do not share space (hunting areas, rest areas, defecation sites, etc.). Domestic cats living in a group therefore have a much higher epidemiological risk of mutation. After this mutation, the FCoV acquires a tropism for macrophages, while losing intestinal tropism.[7]
In a large group of cats, n, the epidemiological risk of mutation (E) is higher and expressed theoretically as: E = n2 −n. A house hosting 2 cats therefore has risk of mutation E = 2. When 4 kittens (6 cats in total) are born into this house, the risk increases from 2 to 30 (62−6). Overcrowding increases the risk of mutation and conversion from FECV to FIPV, which constitutes a major risk factor for the development of feline infectious peritonitis (FIP) cases. FIP has been shown to develop in cats whose immunity is low; such as younger kittens, old cats, immunosuppression due to viral—FIV (feline immunodeficiency virus) and/or FeLV (feline leukemia virus) and stress, including the stress of separation and adoption.[7]
Infection of macrophages by FIPV is responsible for development of a fatal granulomatous vasculitis, or FIP (see granuloma).[7] Development of FIP depends on two factors: virus mutation and low immunity where virus mutation depends on the rate of mutation of FECV to FIPV and the immune status depends on the age, the genetic pool and the stress level. High immune status will be more effective at slowing down the virus.[7]
Molecular biology
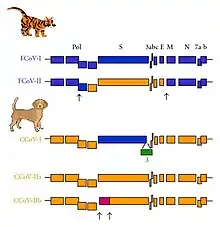
Two forms of feline coronavirus are found in nature: enteric (FECV) and FIP (FIPV). There are also two different serotypes found with different antigens that produce unique antibodies. FCoV serotype I (also called type I) is the most frequent. Type I, that can be defined as 'FECV that could mutate to FIPV type I', is responsible for 80% of the infections. Typically, serotype I FCoV cultures are difficult to perform, with few resulting studies. FCoV serotype II (also called type II) is less frequent and is described as 'FECV type II that can mutate to FIPV type II.' FCoV type II is a recombinant virus type I with spike genes (S protein) replacement from FCoV by the canine coronavirus (CCoV) spikes.[9]
More recent research points to a common ancestor between FCoV and CCoV. This ancestor gradually evolved into FCoV I. An S protein from a yet-unknown virus was passed into the ancestor and gave rise to CCoV, whose S protein was again recombined into FCoV I to form FCoV II. CCoV gradually evolved into TGEV.[10]
FCoV type II
Virus fusion
Coronaviruses are covered with several types of "S proteins" (or E2) forming a crown of protein spikes on the surface of the virus. Coronaviruses take their name from the observation of this crown by electron microscopy. These spikes of Cov (group 1 and serotype II) are responsible for the infection power of the virus by binding the virus particle to a membrane receptor of the host cell—the Feline Amino peptidase N (fAPN).[11][12][13]
The viral receptor: aminopeptidase N (APN)
fAPN (feline), hAPN (human) and pAPN (porcine) differ in some areas of N-glycosylation. All strains of the coronavirus study group 1 (feline, porcine and human) can bind to the feline aminopeptidase N fapn but the human coronavirus can bind to the human APN (HAPN) but not to the porcine type receptor (pAPN) and the pig coronavirus can bind to the porcine APN (pAPN) but not the human type receptor (hAPN). At the cellular level the glycosylation level of enterocytes APN is important for the binding of virus to the receptor.[14][15]
Viral spikes
The FECV spikes have a high affinity for enterocytes fAPN, while the mutant FIPV spikes have a high affinity for the macrophages fAPN. During the viral replication cycle, spikes proteins mature in the host cell Golgi complex with a high mannose glycosylation. This spike manno-glycosylation stage is vital for the acquisition of coronavirus virility.[7][16]
Molecular model of FCoV type I
The receptor
In 2007, it was well established that serotype I did not work with the FCoV fAPN receptor. The FCoV type I receptor still is unknown.[17]
CoV receptors
The human CoV SARS binds to the angiotensin-converting enzyme ACE II. The ACE II is also called L-SIGN (liver/lymph node-specific intracellular adhesion molecules-3 grabbing non-integrin). Coronaviruses bind to macrophages via the Dendritic Cell-Specific Intercellular adhesion molecule-3-Grabbing Non-integrin (DC-SIGN) which is a trans-membrane protein encoded in humans by the CD209 gene.[18] ACE and DC-SIGN are two trans-membrane retrovirus receptors (mannose receptors) which can bind "the plant lectins C-type mannose binding domain".[19]
Aminopeptidase N has the same ability to interact with plant lectins C-type mannose-binding and also serves as a receptor for a retrovirus. Angiotensin-converting enzyme ACE, aminopeptidase A and aminopeptidase N have cascading actions in the renin-angiotensin-aldosterone system, which suggests a common phylogenetic origin between these molecules. Some advanced studies have shown a high homology between the Aminopeptidase N and the Angiotensin-converting enzyme.[20]
Interactions between the viruses and sialic acid
Sialic acid is a component of the complex sugar glycocalix, which is the mucus protecting the gastrointestinal and respiratory mucosa. It is an important facilitating fusion factor of any viruses to its host cell which has been very well studied for flu.
Extensive data also shows that processes using sialic acid are directly involved in the interaction with the receptor's lectins.[21] It has also been demonstrated that swine enteric coronavirus (group 1) fusion to the enterocyte is accomplished via binding to the APN in the presence of the sialic acid.[15][22][23] Feline coronavirus infections are therefore sialic acid dependent.[24][25]
The Porcine epidemic diarrhea virus (PEDV) S protein is 45% identical to FCoV type I spike. An EM structure of it shows sialic acid binding sites. The PEDV receptor is also unknown.[26]
Effects of breast milk on kittens
Colostrum
Other molecules from colostrum and cat milk, could also bear this coverage: lactoferrin, lactoperoxidase, lysozyme, rich proline polypeptide — PRP and alpha-lactalbumine. Lactoferrin has many properties that make it a very good candidate for this anti-coronavirus activity:
- For FCoV group II, it binds to APN.[27]
- For SARS CoV, it binds to ACEs[28]
- It also binds to DC-SIGN of macrophages,[29]
- The lactoferrin anti-viral activity is sialic-acid–dependent.
The structures of the polypeptide chain and carbohydrate moieties of bovine lactoferrin (bLF) are well established. bLF consists of a 689-amino acid polypeptide chain to which complex and high-mannose-type glycans are linked.[30]
Other components
The colostrum and breast milk also contain:
- Many oligosaccharides (glycan) which are known for their anti-viral properties which is thought to be primarily due to their inhibition of pathogen binding to host cell ligands.[31]
- Many maternal immune cells.
- Many cytokines (interferon, etc.), whose role by oro-mucosal route seems very important.[32][33][34]
- Sialic acid: during lactation, neutralizing oligo-saccharides binding sialic acid decreases when it binds increasingly to glycoproteins.[35] (The APN is a glycoprotein.) The anti-viral effect of lactoferrin is increased by the removal of sialic acid.[36]
- Mannan-binding lectins.[37]
Other protective factors
Other assumptions may help to explain this resistance to FCoV infections by kittens. In the first weeks of life, APN could be immature because highly manno-glycosylated.[38] The spikes of CoV could then not be bound. Factors in breastmilk may inhibit the synthesis of fANP by enterocytes, as already described with fructose or sucrose.[39][40][41]
References
- ↑ "ICTV 9th Report (2011) Coronaviridae". International Committee on Taxonomy of Viruses (ICTV). Retrieved 10 January 2019.
- 1 2 3 Taharaguchi, Satoshi; Soma, Takehisa; Hara, Motonobu (2012). "Prevalence of Feline Coronavirus Antibodies in Japanese Domestic Cats during the Past Decade". Journal of Veterinary Medical Science. 74 (10): 1355–8. doi:10.1292/jvms.11-0577. PMID 22673084.
- ↑ Hartmann, Katrin (2005). "Feline infectious peritonitis". Veterinary Clinics of North America: Small Animal Practice. 35 (1): 39–79. doi:10.1016/j.cvsm.2004.10.011. PMC 7114919. PMID 15627627.
- ↑ Pedersen, NC; Perron, M; Bannasch, M (2019). "Efficacy and safety of the nucleoside analog GS-441524 for treatment of cats with naturally occurring feline infectious peritonitis". Journal of Feline Medicine and Surgery. 21 (4): 271–281. doi:10.1177/1098612X19825701. PMC 6435921. PMID 30755068.
- ↑ Addie, Diane D.; McDonald, Mike; Audhuy, Stéphane; Burr, Paul; Hollins, Jonathan; Kovacic, Rémi; Lutz, Hans; Luxton, Zoe; Mazar, Shlomit; Meli, Marina L. (2012). "Quarantine protects Falkland Islands (Malvinas) cats from feline coronavirus infection". Journal of Feline Medicine and Surgery. 14 (2): 171–176. doi:10.1177/1098612X11429644. PMID 22314098. S2CID 4989860.
- ↑ Levy, J.K.; Crawford, P.C.; Lappin, M.R.; Dubovi, E.J.; Levy, M.G.; Alleman, R.; Tucker, S.J.; Clifford, E.L. (2008). "Infectious Diseases of Dogs and Cats on Isabela Island, Galapagos". Journal of Veterinary Internal Medicine. 22 (1): 60–65. doi:10.1111/j.1939-1676.2007.0034.x. PMC 7166416. PMID 18289290. S2CID 23423426.
- 1 2 3 4 5 6 7 8 9 Rottier, Peter J. M.; Nakamura, Kazuya; Schellen, Pepijn; Volders, Haukeline; Haijema, Bert Jan (2005). "Acquisition of Macrophage Tropism during the Pathogenesis of Feline Infectious Peritonitis is Determined by Mutations in the Feline Coronavirus Spike Protein". Journal of Virology. 79 (22): 14122–30. doi:10.1128/JVI.79.22.14122-14130.2005. PMC 1280227. PMID 16254347.
- ↑ Le Poder, Sophie (2011-07-31). "Feline and Canine Coronaviruses: Common Genetic and Pathobiological Features". Advances in Virology. 2011: 609465. doi:10.1155/2011/609465. PMC 3265309. PMID 22312347.
- ↑ Herrewegh, Arnold A. P. M.; Smeenk, Ingrid; Horzinek, Marian C.; Rottier, Peter J. M.; De Groot, Raoul J. (May 1998). "Feline Coronavirus Type II Strains 79-1683 and 79-1146 Originate from a Double Recombination between Feline Coronavirus Type I and Canine Coronavirus". Journal of Virology. 72 (5): 4508–14. doi:10.1128/JVI.72.5.4508-4514.1998. PMC 109693. PMID 9557750.
- ↑ Jaimes, Javier A.; Millet, Jean K.; Stout, Alison E.; André, Nicole M.; Whittaker, Gary R. (10 January 2020). "A Tale of Two Viruses: The Distinct Spike Glycoproteins of Feline Coronaviruses". Viruses. 12 (1): 83. doi:10.3390/v12010083. PMC 7019228. PMID 31936749.
- ↑ Tresnan, Dina B.; Holmes, Kathryn V. (1998). "Feline Aminopeptidase N is a Receptor for All Group I Coronaviruses". In Enjuanes, Luis; Siddell, Stuart G.; Spaan, Willy (eds.). The Effects of Noise on Aquatic Life. Advances in Experimental Medicine and Biology. Vol. 730. pp. 69–75. doi:10.1007/978-1-4615-5331-1_9. ISBN 978-1-4419-7310-8. PMID 9782266.
- ↑ Tresnan, Dina B.; Levis, Robin; Holmes, Kathryn V. (December 1996). "Feline aminopeptidase N serves as a receptor for feline, canine, porcine, and human coronaviruses in serogroup I". Journal of Virology. 70 (12): 8669–74. doi:10.1128/JVI.70.12.8669-8674.1996. PMC 190961. PMID 8970993.
- ↑ Holmes, K. V.; Tresnan, D. B.; Zelus, B. D. (1997). "Virus-Receptor Interactions in the Enteric Tract". In Paul, Prem S.; Francis, David H.; Benfield, David A. (eds.). Mechanisms in the Pathogenesis of Enteric Diseases. Advances in Experimental Medicine and Biology. Vol. 412. pp. 125–33. doi:10.1007/978-1-4899-1828-4_20. ISBN 978-1-4899-1830-7. PMID 9192004.
- ↑ Wentworth, D. E.; Holmes, K. V. (2001). "Molecular Determinants of Species Specificity in the Coronavirus Receptor Aminopeptidase N (CD13): Influence of N-Linked Glycosylation". Journal of Virology. 75 (20): 9741–52. doi:10.1128/JVI.75.20.9741-9752.2001. PMC 114546. PMID 11559807.
- 1 2 Schwegmann-Wessels, Christel; Herrler, Georg (2008). "Identification of Sugar Residues Involved in the Binding of TGEV to Porcine Brush Border Membranes". In Cavanagh, Dave (ed.). SARS- and Other Coronaviruses. Methods in Molecular Biology. Vol. 454. pp. 319–29. doi:10.1007/978-1-59745-181-9_22. ISBN 978-1-58829-867-6. PMC 7122611. PMID 19057868.
- ↑ Regan, A. D.; Whittaker, G. R. (2008). "Utilization of DC-SIGN for Entry of Feline Coronaviruses into Host Cells". Journal of Virology. 82 (23): 11992–6. doi:10.1128/JVI.01094-08. PMC 2583691. PMID 18799586.
- ↑ Dye, C.; Temperton, N.; Siddell, S. G. (2007). "Type I feline coronavirus spike glycoprotein fails to recognize aminopeptidase N as a functional receptor on feline cell lines". Journal of General Virology. 88 (6): 1753–60. doi:10.1099/vir.0.82666-0. PMC 2584236. PMID 17485536.
- ↑ Curtis, Benson M.; Scharnowske, Sonya; Watson, Andrew J. (1992). "Sequence and expression of a membrane-associated C-type lectin that exhibits CD4-independent binding of human immunodeficiency virus envelope glycoprotein gp120". Proceedings of the National Academy of Sciences. 89 (17): 8356–60. Bibcode:1992PNAS...89.8356C. doi:10.1073/pnas.89.17.8356. JSTOR 2361356. PMC 49917. PMID 1518869.
- ↑ Lozach, Pierre-Yves; Burleigh, Laura; Staropoli, Isabelle; Amara, Ali (2007). "The C Type Lectins DC-SIGN and L-SIGN". Glycovirology Protocols. Methods in Molecular Biology. Vol. 379. pp. 51–68. doi:10.1007/978-1-59745-393-6_4. ISBN 978-1-58829-590-3. PMC 7122727. PMID 17502670.
- ↑ Armelle, Armelle; Ascher, P.; Roques, B.-P. (1993). Analyse structurale du site actif de trois métallopeptidases à zinc: Endopeptidase Neutre-24. II, Aminopeptidase N et Enzyme de Conversion de l'Angiotensine [Structural analysis of the active site of three Zinc-metallopeptidases: Neutral Endopeptidase-24.11, Aminopeptidase N and Angiotensin Converting Enzyme] (PhD Thesis) (in French). Paris: Université de Paris. p. 160. OCLC 490188569. INIST 163816.
- ↑ Lehmann, F.; Tiralongo, E.; Tiralongo, J. (2006). "Sialic acid-specific lectins: Occurrence, specificity and function". Cellular and Molecular Life Sciences. 63 (12): 1331–54. doi:10.1007/s00018-005-5589-y. PMC 7079783. PMID 16596337.
- ↑ Schwegmann-Wessels, C.; Zimmer, G.; Laude, H.; Enjuanes, L.; Herrler, G. (2002). "Binding of Transmissible Gastroenteritis Coronavirus to Cell Surface Sialoglycoproteins". Journal of Virology. 76 (12): 6037–43. doi:10.1128/JVI.76.12.6037-6043.2002. PMC 136196. PMID 12021336.
- ↑ Schwegmann-Wessels, C.; Zimmer, G.; Schröder, B.; Breves, G.; Herrler, G. (2003). "Binding of Transmissible Gastroenteritis Coronavirus to Brush Border Membrane Sialoglycoproteins". Journal of Virology. 77 (21): 11846–8. doi:10.1128/JVI.77.21.11846-11848.2003. PMC 229351. PMID 14557669.
- ↑ Paltrinieri, Saverio; Gelain, Maria E.; Ceciliani, Fabrizio; Ribera, Alba M.; Battilani, Mara (2008). "Association between faecal shedding of feline coronavirus and serum α1-acid glycoprotein sialylation". Journal of Feline Medicine & Surgery. 10 (5): 514–8. doi:10.1016/j.jfms.2008.04.004. PMC 7129531. PMID 18701332.
- ↑ Paltrinieri, S; Metzger, C; Battilani, M; Pocacqua, V; Gelain, M; Giordano, A (2007). "Serum α1-acid glycoprotein (AGP) concentration in non-symptomatic cats with feline coronavirus (FCoV) infection". Journal of Feline Medicine & Surgery. 9 (4): 271–7. doi:10.1016/j.jfms.2007.01.002. PMC 7129318. PMID 17344083.
- ↑ Wrapp, Daniel; McLellan, Jason S.; Gallagher, Tom (13 November 2019). "The 3.1-Angstrom Cryo-electron Microscopy Structure of the Porcine Epidemic Diarrhea Virus Spike Protein in the Prefusion Conformation". Journal of Virology. 93 (23). doi:10.1128/JVI.00923-19. PMC 6854500. PMID 31534041.
- ↑ Ziere GJ, Kruijt JK, Bijsterbosch MK, Berkel TJ (June 1996). "Recognition of lactoferrin and aminopeptidase M-modified lactoferrin by the liver: involvement of the remnant receptor". Zeitschrift für Gastroenterologie. 34 (3): 118–21. PMID 8767485.
- ↑ Centeno, José M.; Burguete, María C.; Castelló-Ruiz, María; Enrique, María; Vallés, Salvador; Salom, Juan B.; Torregrosa, Germán; Marcos, José F.; Alborch, Enrique; Manzanares, Paloma (2006). "Lactoferricin-Related Peptides with Inhibitory Effects on ACE-Dependent Vasoconstriction". Journal of Agricultural and Food Chemistry. 54 (15): 5323–9. doi:10.1021/jf060482j. PMID 16848512.
- ↑ Groot, F.; Geijtenbeek, T. B. H.; Sanders, R. W.; Baldwin, C. E.; Sanchez-Hernandez, M.; Floris, R.; Van Kooyk, Y.; De Jong, E. C.; Berkhout, B. (2005). "Lactoferrin Prevents Dendritic Cell-Mediated Human Immunodeficiency Virus Type 1 Transmission by Blocking the DC-SIGN--gp120 Interaction". Journal of Virology. 79 (5): 3009–15. doi:10.1128/JVI.79.5.3009-3015.2005. PMC 548463. PMID 15709021.
- ↑ Pierce, Annick; Colavizza, Didier; Benaissa, Monique; Maes, Pierrette; Tartar, Andre; Montreuil, Jean; Spik, Genevieve (1991). "Molecular cloning and sequence analysis of bovine lactotransferrin". European Journal of Biochemistry. 196 (1): 177–84. doi:10.1111/j.1432-1033.1991.tb15801.x. PMID 2001696.
- ↑ Newburg, David S.; Ruiz-Palacios, Guillermo M.; Morrow, Ardythe L. (2005). "Human Milk Glycans Protect Infants Against Enteric Pathogens". Annual Review of Nutrition. 25: 37–58. doi:10.1146/annurev.nutr.25.050304.092553. PMID 16011458.
- ↑ Dec M, Puchalski A (2008). "Use of oromucosally administered interferon-alpha in the prevention and treatment of animal diseases". Polish Journal of Veterinary Sciences. 11 (2): 175–86. PMID 18683548.
- ↑ Tovey, Michael G. (June 2002). "Special Oromucosal Cytokine Therapy : Mechanism(s) of Action". The Korean Journal of Hepatology. 8 (2): 125–31. PMID 12499797.
- ↑ Schellekens, Huub; Geelen, Gerard; Meritet, Jean-François; Maury, Chantal; Tovey, Michael G. (2001). "Oromucosal Interferon Therapy: Relationship Between Antiviral Activity and Viral Load". Journal of Interferon & Cytokine Research. 21 (8): 575–81. doi:10.1089/10799900152547830. PMID 11559435.
- ↑ Martín, M.-J.; Martín-Sosa, S.; García-Pardo, L.-A.; Hueso, P. (2001). "Distribution of Bovine Milk Sialoglycoconjugates During Lactation". Journal of Dairy Science. 84 (5): 995–1000. doi:10.3168/jds.S0022-0302(01)74558-4. PMID 11384055.
- ↑ Superti, Fabiana; Siciliano, Rosa; Rega, Barbara; Giansanti, Francesco; Valenti, Piera; Antonini, Giovanni (2001). "Involvement of bovine lactoferrin metal saturation, sialic acid and protein fragments in the inhibition of rotavirus infection". Biochimica et Biophysica Acta (BBA) - General Subjects. 1528 (2–3): 107–15. doi:10.1016/S0304-4165(01)00178-7. hdl:11573/83235. PMID 11687297.
- ↑ Trégoat, Virginie; Montagne, Paul; Béné, Marie-Christine; Faure, Gilbert (2002). "Changes in the mannan binding lectin (MBL) concentration in human milk during lactation". Journal of Clinical Laboratory Analysis. 16 (6): 304–7. doi:10.1002/jcla.10055. PMC 6807810. PMID 12424804.
- ↑ Danielsen, E.Michael; Hansen, Gert H.; Niels-Christiansen, Lise-Lotte (1995). "Localization and biosynthesis of aminopeptidase N in pig fetal small intestine". Gastroenterology. 109 (4): 1039–50. doi:10.1016/0016-5085(95)90561-8. PMID 7557068.
- ↑ Danielsen, E. Michael (1992). "Folding of intestinal brush border enzymes. Evidence that high-mannose glycosylation is an essential early event". Biochemistry. 31 (8): 2266–72. doi:10.1021/bi00123a008. PMID 1347233.
- ↑ Danielsen, E. Michael; Hansen, Gert H.; Wetterberg, Lise-Lotte (December 1991). "Morphological and functional changes in the enterocyte induced by fructose". The Biochemical Journal. 280 (2): 483–9. doi:10.1042/bj2800483. PMC 1130574. PMID 1684104.
- ↑ Danielsen, E. Michael (August 1989). "Post-translational suppression of expression of intestinal brush border enzymes by fructose". The Journal of Biological Chemistry. 264 (23): 13726–9. doi:10.1016/S0021-9258(18)80059-X. PMID 2569463.
External links
- Dr ADDIE website focused research about FIP
- Coronavirus Site général
- Coronavirus site général
- Coronavirus Pictures
Higher taxonomy: Riboviria > Orthornavirae > Pisuviricota > Pisoniviricetes > Nidovirales > Cornidovirineae > Coronaviridae | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||