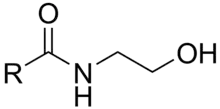
An N-acylethanolamine (NAE) is a type of fatty acid amide where one of several types of acyl groups is linked to the nitrogen atom of ethanolamine, and highly metabolic formed by intake of essential fatty acids through diet by 20:4, n-6 and 22:6, n-3 fatty acids,[1][2] and when the body is physically and psychologically active,.[3][4] The endocannabinoid signaling system (ECS) is the major pathway by which NAEs exerts its physiological effects in animal cells with similarities in plants, and the metabolism of NAEs is an integral part of the ECS,[5] a very ancient signaling system, being clearly present from the divergence of the protostomian/deuterostomian,[6][7] and even further back in time, to the very beginning of bacteria, the oldest organisms on Earth known to express phosphatidylethanolamine, the precursor to endocannabinoids, in their cytoplasmic membranes. Fatty acid metabolites with affinity for CB receptors are produced by cyanobacteria, which diverged from eukaryotes at least 2000 million years ago (MYA), by brown algae which diverged about 1500 MYA, by sponges, which diverged from eumetazoans about 930 MYA, and a lineages that predate the evolution of CB receptors, as CB1 – CB2 duplication event may have occurred prior to the lophotrochozoan-deuterostome divergence 590 MYA. Fatty acid amide hydrolase (FAAH) evolved relatively recently, either after the evolution of fish 400 MYA, or after the appearance of mammals 300 MYA, but after the appearance of vertebrates. Linking FAAH, vanilloid receptors (VR1) and anandamide (NAE 20:4) implies a coupling among the remaining ‘‘older’’ parts of the endocannabinoid system, monoglyceride lipase (MGL), CB receptors, that evolved prior to the metazoan-bilaterian divergence (ie, between extant Hydra and leech), but were secondarily lost in the Ecdysozoa, and 2-Arachidonoylglycerol (2-AG).[8]
These amides conceptually can be formed from a fatty acid and ethanolamine with the release of a molecule of water, but the known biological synthesis uses a specific phospholipase D to cleave the phospholipid unit from N-acylphosphatidylethanolamines.[9] Another route relies on the transesterification of acyl groups from phosphatidylcholine by an N-acyltransferase (NAT) activity. The suffixes -amine and -amide in these names each refer to the single nitrogen atom of ethanolamine that links the compound together: it is termed "amine" in ethanolamine because it is considered as a free terminal nitrogen in that subunit, while it is termed "amide" when it is considered in association with the adjacent carbonyl group of the acyl subunit. Names for these compounds may be encountered with either "amide" or "amine" varying by author.[10]
N-acylethanolamines (NAEs) are broken down, or hydrolysed, by fatty acid amide hydrolase (FAAH) to ethanolamine (MEA) and their corresponding fatty acid, arachidonic acid. FAAH is activated during stress exposure circumstances, which also raises the neuronal excitability in the amygdala, a critical brain area that mediates anxiety, and the anxiolytic outcome of CB1 receptor activation.[11] Inhibition of FAAH has been shown to increase the levels of NAEs in vivo and to produce desirable phenotypes, that produce analgesic, anxiolytic, neuroprotective, and anti-inflammatory effects,[12] like in high-level performance athletes (i.e., elite athletes) that present an extraordinary interindividual variability of physical, but also mental traits, that greatly influence their sports accomplishments and their career longevity, by an FAAH genetic polymorphism that produce the SNP rs324420 (C385A allele), associated with a higher sensitivity of FAAH to proteolytic degradation and a shorter half-life, as compared to the C variant, as the A variant displays normal catalytic properties, but an enhanced sensitivity to degradation, leading to increased NAE and anandamide (AEA) signaling.[11] Activation of the cannabinoid receptor CB1 or CB2 in different tissues, including skin, inhibit FAAH, and thereby increases endocannabinoid levels.[13]
Examples
Examples of N-acylethanolamines include:[14]
- Anandamide (N-arachidonoylethanolamine; NAE) or arachidonoylethanolamine (AEA: C22H37NO2; 20:4, ω-6) is the amide of arachidonic acid (C20H32O2; 20:4, ω-6) and ethanolamine (MEA: C2H7NO).[15] It is the ligand of both cannabinoid receptors and vanilloid receptor that attenuates pain sensation.[16][17][18][19] Where binding to cannabinoid receptors is coupled to nitric oxide (NO) release in the central nervous system of invertebrates and in peripheral immune cells of both invertebrates and humans.[20]
- N-Palmitoylethanolamine (PEA: C18H37NO2; 16:0) is the amide of palmitic acid (C16H32O2; 16:0) and ethanolamine. It is a ligand at CB2 receptors,[21][22] and PPAR-α.[23] It has anti-inflammatory activity and also attenuates pain sensation in mammals, whose levels are increased following neuroinflammatory or neuropathic conditions in both animals and humans, possibly to exert a local anti-inflammatory and analgesic action.[19][24][25][26] NAE 16:0 has also been identified in plants including corn, and seeds of cotton, okra, tomato, castor bean, soya bean and peanuts,[27] but its physiological functions remain unknown,[28]
- N-alpha-Linoleoylethanolamide (ALEA: C20H35NO2; 18:3, ω-3) or Anandamide (18:3, n-3),[29] is a metabolic product of the omega-3 fat α-linoleic acid (ALA: C18H30O2; 18:3, ω-3) and ethanolamine (MEA: C2H7NO).[30]
- N-Oleoylethanolamine (OEA: C20H39NO2; 18:1, ω-9) is the amide of oleic acid (C18H34O2; 18:1) and ethanolamine. It has anorexic effects and enables fat breakdown by stimulating PPAR-alpha.[31] In plants, NAE 18:1 is present abundantly in dry seeds and levels decline during seed imbibition,[27][32] but its physiological functions are yet to be elucidated. In humans, plasma OEA levels are also found positively correlated with positive mood and emotions.[4] OEA acting mostly at peroxisome proliferator‐activated receptor‐α (PPAR‐α) nuclear receptors and, to some extent, TRPV1 channels.[33]
- N-Stearoylethanolamine (SEA: C20H41NO2; 18:0) is the amide of stearic acid (C18H36O2) and ethanolamine (MEA: C2H7NO). It has pro-apoptotic activity. It operates independently of the known cannabinoid and vanilloid receptors targeted by anandamide.[31] It is an inhibitor of the sphingolipid signaling pathway, via specific ceramidase inhibition (ceramidase converts ceramide to sphingosine) and blocks the effects of TNF- and arachidonic acid on intracellular Ca2+ concentration.[34][35][36]
- N-Linoleoyl ethanolamide (MEA: C20H37NO2; 18:2, ω-6) or Anandamide (18:2, n-6) is the ethanolamide of linoleic acid (LA: C18H32O2; 18:2, ω-6) and its metabolized incorporated ethanolamine (MEA: C2H7NO),[37] is the first natural inhibitor of fatty acid amide hydrolase (FAAH) discovered, also derives from hydroperoxides of NAE 20:4 (AEA) or its linoleoyl analogues by lipoxygenase action.[20]
- N-Docosahexaenoyl ethanolamine (DHEA: C24H37NO2; 22:6, ω-3), or Anandamide (22:6, n-3) "synaptamide", is the non-oxidative produced amide of the mainly fish oil derived, docosahexaenoic acid (DHA: C22H32O2; 22:6, ω-3) and ethanolamine (MEA: C2H7NO).[38] It can act at CB1 and CB2 receptors,[39][40] and have anti-proliferative effects on prostate cancer cell lines and promotes synaptogenesis, neurogenesis and neuritogenesis,[41][42] and as an endogenous metabolite of DHA, it promotes brain development and function.[43]
- N-Docosatetraenoylethanolamine (DEA: C24H41NO2; 22:4,ω-6) act on the CB1 receptor,[21] and possible CB2.[34]
- N-Eicosapentaenoylethanolamide (EPEA: C22H35NO2; 20:5, ω-3) or Anandamide (20:5, n-3). It is the amide of eicosapentaenoic acid (EPA: C20H30O2; 20:5, n-3),[44] that can act on CB1 and CB2 receptors in combination with PPAR-gamma to decrease LPS induced adipocyte IL-6 and MCP-1 levels.[39][40]
- N-homo-gamma-linolenoylethanolamine, or Anandamide (20:3,n-6) (HGLEA: C22H39NO2; 20:3,ω-6).[45]
These bioactive lipid amides are generated by the membrane enzyme NAPE-PLD, and natural bile acids regulate this essential process.[46] An in vivo active NAPE-PLD inhibitor called LEI-401 was found to be CNS-active and modulated NAE biosynthesis. It had similar effects as a cannabinoid CB1 receptor antagonist, which could be reversed by co-treatment with a FAAH inhibitor.[47]
At least two pathways distinct from NAPE-PLD have been proposed as metabolic pathways for NAE 20:4 (AEA) formation. One pathway involves the double-O-deacylation of NAPEs by α,β-hydrolase (ABHD4) to form glycerophospho-N-acylethanolamines (GP-NAEs),[48] followed by conversion of these intermediates to NAEs by glycerophosphodiesterase-1 (GDE1). Another pathway utilizes a phospholipase C (PLC) to produce phopho-N-arachidonoylethanolamine (pAEA) from NAPE, widely found in phospholipids,[49] followed by conversion of pAEA into NAE 20:4 (AEA) by phosphatases such as PTPN22 and SHIP1.[50]
The effects of NAE 20:4 (AEA) and another endocannabinoid 2-Arachidonoylglycerol (2-AG: C23H38O4; 20:4, ω-6), with tissue levels of 2-AG usually several tens to several hundreds of times those of AEA,[51] is found to be enhanced by “entourage compounds”, NAEs that inhibit their hydrolysis via substrate competition, and thereby prolong their action. These compounds include N-palmitylethanolamide (PEA, NAE 16:0), N-oleoylethanolamide (SEA, NAE 18:0), and cis-9-octadecenoamide (OEA, oleamide, NAE 18:1).[52]
All are members of the endocannabinoidome, a complex lipid signaling system composed of more than 100 of fatty acid-derived mediators and their receptors, its anabolic and catabolic enzymes of more than 50 proteins, which are deeply involved in the control of energy metabolism and its pathological deviations,[53] as well as immunosuppression,[54] and some NAE members, do not activate the CB1 and CB2 receptors efciently, but instead activate other receptors (e.g. peroxisome proliferator-activated receptors (PPAR)-α/γ, G-protein coupled receptors (GPR) 55, 110, 118, 119, TRPV1 channels), known to counteract metabolic disorders in animal models, by gut bacterial families (e.g. Veillonellaceae, Peptostreptococcaceae and Akkermansiaceae) associated with variations in most NAEs and omega3-derived 2-monoacylglycerols (2‑MAGs), congeners of 2-AG, as gut microbiota communities and the host endocannabinoidome also seem to be interrelated in a mutual crosstalk controlling whole body metabolism,[55][56] and onset and development of chronic intestinal inflammation.[57]
NAE's are also involved in modulation of different physiological processes such as pain, stress, anxiety, appetite, cardiovascular function and inflammation. A study suggests the presence of an endogenous NAE tone control emotional behavior.[47]
Raphael Mechoulam that described and named Anandamide in 1992. He said:
Look, I believe there are 8 billion people on this planet, and I believe there are 8 billion different personalities. One way of explaining it is, there are several hundred compounds, endocannabinoid-like compounds. They are like anandamide in their chemical structure, that are present in the brain, and it is quite possible that each one of us, has a different, slightly different level of these compounds. And it is quite possible that differences in the endocannabinoid system, endocannabinoid-like system, can have something to do with the different personalities, and that ratios of 10 of these to 10 of others and so on will cause that.[16] in the YouTube video The Scientist, released in 2015.[58]
Beyond vertebrates NAEs are also found to have signaling roles in more primitive organism, implicated as metabolic signals that coordinate nutrient status and lifespan determination in Caenorhabditis elegans, and detected in organisms as diverse as yeast (Saccharomyces cerevisiae), freshwater fish (Esox lucius and Cyprinus carpio), bivalve mollusc (Mytilus galloprovincialis), protists (Tetrahymena thermophila), slime mold (Dictyostelium discoideum), microbes such as bacteria, fungi, and viruses, are all organisms that appear to regulate their endogenous NAE levels via similar enzymatic machinery as mammalian vertebrates, show a widespread occurrence of NAEs, from single-celled organisms to humans, and a highly conserved role for this group of lipids in cell signaling.[59][22] Studies in amphibians and birds show that endocannabinoid signaling may function as a behavioral switch, allowing redirection from less- to more-essential behaviors in response to emergent environmental changes,[60][61] and provide evidence of cannabinoid modulation of aggression, emesis, feeding behavior, locomotor activity, reproductive behaviors, vocal learning, sensory perception and stress responses.[62][63]
NAE and the reproductive system
Several researchers have found, that NAE, and especially 20:4 anandamide (AEA: C22H37NO2; 20:4, ω-6), is a part of the reproductive system,[64] and play a fundamental role for a healthy and successful pregnancy.
A 2006 report from the Pediatrics Department at Vanderbilt University characterized NAE 20:4 (AEA) as “an emerging concept in female reproduction”, because they found a “cannabinoid sensor” mechanism to influence several crucial steps during early pregnancy.
The Vanderbilt research team termed this “endocannabinoid signaling in preimplantation embryo development and activation”, because one of the first things the fertilized embryo must do, is to attach itself to the lining of the uterus, and without becoming attached to the uterine wall, which forms the umbilical cord, there will be no pregnancy. NAE 20:4 (AEA) plays a key role, because, for the embryo to become attached to the lining of the uterus, a particular amount (temporary reduction by high Fatty acid amide hydrolase FAAH) of NAE 20:4 (AEA),[65] present at the uterine lining (uterine epithelium), is necessary for the fertilized embryo can attach itself to the uterine wall, i.e. implantation. NAE 20:4 (AEA) uses the CB1 receptors, that are at high levels on the blastocyst (fertilized egg), to this attachment. So, the amount of NAE 20:4 (AEA) directs the outcome of the attachment to the uterine wall via CB1, and thereby, the outcome to pregnancy,[66][67] by synchronizing trophoblast differentiation and uterine preparation to the receptive state.
However, low FAAH expression and high NAE 20:4 (AEA) levels at the interimplantation sites, prior to successful implantation, have been reported, and a later possible miscarriage, as AEA levels are inversely correlated with FAAH levels in peripheral blood mononuclear cells (PMNCs) and FAAH levels are found lower in women who consequently miscarry compared with those who progress beyond the first trimester. A consequence also found in women undergoing in vitro fertilization and embryo transfer, as low activity of FAAH in PMNCs and high plasma AEA levels after embryo transfer show failure to achieve a successful pregnancy. As well, high AEA level also inhibits BeWo trophoblast cell proliferation, in a dose-dependent manner, via the CB2 receptor, suggest that FAAH acts as a barrier to the AEA maternal-fetal transfer. So, high plasma AEA levels can be used as a marker of early pregnancy loss in patients with threatened miscarriage, as altered modulation of the ECS contribute to the spontaneous pregnancy loss.[68]
This is in line with a study of 50 women, where NAE 20:4 (AEA) hydrolase activity was lower in the seven women who miscarried than in the 43 who did not (60.43 pmol/min per mg protein [SD 29.34] vs 169.60 pmol/min per mg protein [30.20], and another study showing that all 15 women in the low AEA hydrolase group had miscarriages, compared with one of the 105 women with high concentrations at or above the threshold of hydrolase.[69]
An earlier 2004 research into the course of ectopic pregnancy, a result of embryo retention in the fallopian tube, found that decoupled cannabinoid receptor CB1, can cause retention of embryos in the mouse oviduct, and lead to pregnancy failure, as either silencing or amplification of NAE 20:4 (AEA) signaling via CB1 receptors causes oviductal retention or blastocyst incompetence for implantation. The report estimates that aberrant cannabinoid signaling impedes coordinated oviductal smooth muscle contraction and relaxation, which are crucial to normal oviductal embryo transport. This was also seen in wild-type mice treated with methanandamide (AM-356; C23H39NO2, 20:4, n6), and thereby concluded, that a colocalization of CB1 in the oviduct muscularis implicate a basal endocannabinoid tone of NAE 20:4 (AEA) is needed for oviductal motility and for normal journey of embryos into the uterus.[70]
Another 2004 study published in the American Journal of Obstetrics and Gynecology find NAE 20:4 (AEA) and the phytocannabinoid Δ9-tetrahydrocannabinol (THC: C21H30O2), that can mimic NAE 20:4 (AEA) by direct activation at CB1, as the CB1 carboxyl-terminus have critical structures important for CB1 activity and regulation in the receptor life cycle including activation, desensitization, and internalization,[71] the CB2 receptors,[72][73] and markedly lowering AEA content levels,[74] enhance the function of glycine receptors (GlyRs),[75] activate the nuclear receptor family, peroxisome proliferator-activated receptors (PPARs),[76] able to switch from agonist to antagonist depending on firing rate,[77] and to protect neonatal cardiac cells against hypoxia via CB2 receptor activation by induction of NO production,[78][79] to exert a direct relaxant effect on human pregnant myometrium in vitro, with equal potency for both compounds, which was mediated through the CB1 receptor. This means that the middle layer of the uterine wall, where also CB2 reseptors are expressed, is modulated by NAE 20:4 (AEA) as well.[80]
Likewise, there is also demonstrated CB1 expression in the first trimester placenta characterized by a spatial-temporal modulation. But, at term, there is found lack of FAAH and high CB1 expression at placental villous tissue of non-laboring compared with laboring.[68]
After birth, CB1 receptors appears to be critical for milk sucking by newborn, as it apparently activate oral-motor musculature, by 2-AG (C23H38O4; 20:4 ω-6) in the breast milk, activation, as elevated levels of 2-AG modulate infant appetite and health,[81] as well as NAE 20:4 (AEA) act as a neuroprotectant, also by providing retrograde signaling in the developing postnatal brain, with observations suggest that children may be less prone to psychoactive side effects of Δ9-tetrahydrocannabinol (THC: C21H30O2) or endocannabinoids than adults, as very low density of CB1, and neonatal cardiac cells express CB2, but not CB1 receptors,[79] suggest a promising future for cannabinoids in pediatric medicine for conditions including non-organic failure-to-thrive and cystic fibrosis.[67][82]
Mood
As the euphoric feeling described after running, called the "runners high" is, at least in part, due to increased circulating endocannabinoids (eCBs), and these lipid signaling molecules are involved in reward, appetite, mood, memory and neuroprotection, an analysis of endocannabinoid concentrations and moods after singing, dancing, exercise and reading in healthy volunteers, showed that singing increased plasma levels of anandamide (AEA) by 42%, palmitoylethanolamine (PEA) by 53% and oleoylethanolamine (OEA) by 34%, and improved positive mood and emotions. Dancing did not affect eCB levels but decreased negative mood and emotions. Cycling increased OEA levels by 26% and reading increased OEA levels by 28%. All the ethanolamines were positively correlated with heart rate. As so, the plasma OEA levels were positively correlated with positive mood and emotions, and AEA levels were seen positively correlated with satiation.[4]
However in posttraumatic stress disorder (PTSD), circulating NAE 20:4 (AEA) are found associated with overall mood states and exercise-induced improvements in women with and without PTSD, as AEA significantly increased following aerobic exercise for both groups, whereas the circulation of the endocannabinoid 2-AG only increased in women without PTSD, thereby AEA was associated with lower depressive mood, confusion, and total mood disturbance within the PTSD group and consistent with the discovery of a greater eCB tone, and particularly AEA, following pharmacological and/or non-pharmacological manipulations that may be beneficial for improving psychological outcomes, as mood and cognition among PTSD and possibly other psychiatric populations.[83]
NAE and endocannabinoids is an integral component of stress recovery, both centrally and peripherally, through regulation of the HPA axis, and reduction in circulating NAE 20:4 (AEA) content in major depression, and exposure to stress, is found to increase inflammatory markers by down-regulating the circulating content of the endogenous anti-inflammatory molecules, through their activation of PPAR-α, palmitoylethanolamine (PEA: C18H37NO2; 16:0) and Oleoylethanolamine (OEA: C20H39NO2; 18:1, ω-9), as NAE catabolism is accelerated by stress and by the same FAAH catabolic pathway.[84]
Administration of CB1 receptor antagonists to humans has been found to increase indices of depression and anxiety. Accordingly, the deficit in circulating endocannabinoids and activation of CB1, documented in individuals with major depression, may contribute to the emotional sequelae associated with this disease. And it is therefore speculated, that this reduction in circulating endocannabinoid and NAE content in depression may be associated with the increased rates of inflammation, cardiovascular disease and autoimmune dysfunction seen in this disease.[84] And in other stress-associated psychiatric disorders, like posttraumatic stress disorder (PTSD) and borderline personality disorder (BPD), characterized by intense and rapidly changing mood states as well as chronic feelings of emptiness, impulsivity, fear of abandonment, unstable relationships, and unstable self-image, showing significantly and cronically reduced content of the NAE 20:4 (AEA) that attenuate depressive and anxious symptoms, by elevated fatty acid amide hydrolase (FAAH) in the amygdala-prefrontal cortex (PFC), that subserves emotion regulation and used to measures of hostility and aggression, provide preliminary evidence of elevated FAAH binding in PFC in any psychiatric condition,[85] may be of great therapeutic interest to psychiatry.[86][87] And consistent with the model that lower endocannabinoid tone could perturb PFC circuitry that regulates emotion and aggression,[88] and the feeling of loneliness, as social contact increases, whereas isolation decreases, the production of the endogenous marijuanna-like neurotransmitter, NAE 20:4 (AEA) in nucleus accumbens (NAc), where activation of CB1 are necessary and sufficient to express the rewarding properties of social interactions, i.e. social contact reward.[89]
And possible why cannabinoids are seen highly used in the prison population, and among those who have been imprisoned, and is clearly involved in daily life in prison, where detainees in some prisons estimated the current use of cannabis/hashish to be as high as 80%, and staff estimate 50%, described analgesic, calming, self-help to go through the prison experience, relieve stress, facilitate sleep, prevent violence, and a social peacemaker, where the introduction of a more restrictive regulation induced fear of violence, increased trafficking and a shift to other drug use.[90][91] As seen in the Danish prisons that reflect a ‘treatment guarantee’ embedded in a policy of zero tolerance and intensified disciplinary sanctions, launched by the Danish Government (Regeringen, 2003) and inspired by US drug policy called The Fight against Drugs, with introduction of better fence systems, more sniffer-dogs, and cell and body search of inmates, with an increasingly repressive response to drugs, including zero tolerance and harsher punishment like isolation, that reflects the same chain of destruction as the steps seen to the Holocaust,[92] to make clients drug free and preparing them for a life without crime.[93]
Released to a daily life environment, where the highest scores for quality of life is observed among habitual cannabis users, followed by occasional users, whereas both non-users and dysfunctional users present less favorable score, and non-users reported more depression or anxiety symptoms and a lower quality of life, than occasional and habitual users, found in a Brazilian cross-sectional study involving more than 7400 adults (6620 recreational cannabis users and 785 non-users), even illegal.[94][95] And other findings imply that a causal link between marijuana use and violence is primarily due to its illegality, and thus would not exist in an environment in which marijuana use, at least medicinally (MML), as a first choice in any situation, is legalized, to correct the injustices of cannabis prohibition, as the legalization of cannabis for adult use is found being increasingly embraced in several countries and local entities, coursed by the economic and human suffering of cannabis prohibition, which have fallen most heavily upon disadvantaged minority populations, and for countries, in which cannabis consumption before constituted a traditional habit, also in religion, and practiced for hundreds or thousands of years,[96][97][98] without being subject to any social opprobrium, as no correlation between Marijuana use and criminal behavior are found, correlates with a reduction in homicide and assault rates, after introduction of state MML.[99][100][101] Followed by an almost 5% estimated reduction in the total suicide rate, for the period 1990 through 2007, with an 11% percent reduction for 20- through 29-year-old males, and a 9% reduction in the suicide rate of 30- through 39-year-old males.[102] And the secondary mortality attributing to herbal cannabis is found extremely rare, and usually associated with misadventures with law enforcement, and the prison experience and of solitary confinements.[103][104]
Longevity
A study of 42 eighty years old (octogenarians) humans living in the east-central mountain area of Sardinia, a High-Longevity Zone (HLZ) in Italy, have found, that the endocannabinoidome related circulating NAEs and familiar fatty acids are associated with a longer human life or longevity, as increased conjugated linoleic acid (CLA: C18H32O2; 18:2, n-6) and heptadecanoic acid (C17H34O2; 17:0), elevated palmitoleic acid (POA; C16H30O2; 16:1, n-7), a conjugate acid of a palmitoleate (C16H29O2; 16:1, n-7),[105] where n-7 fatty acids are precursors for the production of omega-4 fatty acids like palmitolinoleic acid (16:2),[106] and a significantly increased level of NAE 22:6 (DHEA: C24H37NO2; 22:6, n-3), the metabolite of DHA (C22H32O2; 22:6, n-3), and the two endocannabinoids NAE 20:4 (AEA: C22H37NO2; 20:4, ω-6) and 2-arachidonoyl-glycerol (2-AG: C23H38O4; 20:4, n-6), as well of increased NAE 18:1 (OEA: C20H39NO2; 18:1, ω-9), the amide of palmitic acid (C16H32O2; 16:0) and ethanolamine (MEA: C2H7NO), and increase of 2-linoleoyl-glycerol (2-LG; C21H38O4; 18:2, n-6),[107] derived from linoleic acid (LA: C18H32O2; 18:2, n-6), can indicate a metabolic pattern potentially protective from adverse chronic conditions, and show a suitable physiological metabolic pattern, that may counteract the adverse stimuli leading to age-related disorders such as neurodegenerative and metabolic diseases.[108]
It is found that 3T3-L1 adipocytes convert eicosapentaenoic acid (EPA: C20H30O2; 20:5, ω-3) to NAE 20:5 (EPEA: C22H35NO2; 20:5, ω-3) or Anandamide (20:5, n-3) and docosahexaenoic acid (DHA: C22H32O2; 22:6, ω-3) to NAE 22:6 (DHEA: C24H37NO2; 22:6, ω-3), or Anandamide (22:6, n-3). This conversion to EPEA and DHEA decrease IL-6 and MCP-1 levels, and the combined incubations with PPAR-gamma and CB2 antagonists, suggest a role of these receptors in mediating the reduction of IL-6 by DHEA. These results are in line with the hypothesis, that in addition to other pathways, this formation of NAEs may contribute to the biological activity of n-3 PUFAs, and different targets, including the endocannabinoid system, may be involved in the immune-modulating activity of fish-oil derived NAEs.[39]
The importance of a low ratio of omega-6 to omega-3 essential fatty acids
Studies have found that humans evolved on a diet with a ratio of omega-6 (n-6) to omega-3 (n-3) essential fatty acids (EFA) of about 1:1, whereas in today's Western diets the ratio is 15/1–16.7/1, or even more. The excessive amounts of n-6 polyunsaturated fatty acids (PUFA) and a very high n-6/n-3 ratio, promote the pathogenesis of many diseases, including cardiovascular disease, cancer, and inflammatory and autoimmune diseases, whereas a low n-6/n-3 ratio exert suppressive effects. However, it is found impotent, that this low ratio, should change a bit, depending on disease, as the ratio of 2.5/1 reduce rectal cell proliferation in patients with colorectal cancer, and 2–3/1 suppress inflammation in patients with rheumatoid arthritis, 4/1 is optimum for prevention of cardiovascular disease, showing a 70% decrease in total mortality, and 5/1 have a beneficial effect on patients with asthma, whereas 10/1 have adverse consequences, indicate, that the optimal ‘low ratio’, may vary with the specific disease.[109][60]
The World Health Organization (WHO) estimate hemp, a culture CO2 negative, - a crop that is capable in the carbon cycle of removing more CO2 from the ambient than it emits, where production of biomass produce between 8 and 12 tons of CO2, but seize between 10 and 15 tons per hectare, with the possibility to sequester up to 22 tons of CO2 from the increased dry matter of the stem, where 80% of atmospheric carbon is sequestered and stored, by a nitrogen fertilization between 0 and 120 kg per hectare,[60] with roots that by various physicians and herbalists in the latter part of 17th century, was recommended to treat fever, inflammation, gout, arthritis, and joint pain, as well as skin burns and hard tumors, beside more,[110] as well as to have modest antimicrobial activity against Cryptococcus neoformans by ergost-5-en-3-ol,[111] and potent antimicrobial activity against Escherichia coli by p-coumaroyltyramine,[112][113] as having what is considered to be an optimal 3:1 balance of omega 6 to omega 3 essential fatty acids, and where hempseed oil, of which 80% are polyunsaturated fatty acids, of which 60% are omega-6 linoleic acid (LA: C18H32O2), the precursor of NAE 20:4 (AEA) and other NAEs, and 20% are omega-3 alpha-linolenic acid (ALA: C18H30O2), the precursor of NAE 18:3 (ALEA: C20H35NO2; 18:3, ω-3) or Anandamide (18:3, n-3),[29][30] is the only one that is in perfect balance according to what the human body needs – 3:1, and a pound (454 gram) of hemp seed, of which 43% are protein, can provide all the protein, essential fatty acids, and dietary fiber necessary for human survival for two weeks, or 33 gram a day.[2][114][115][116][117][118]
And their absence has been found responsible for the development of a wide range of diseases such as metabolic disorders,[3] cardiovascular disorders, inflammatory processes, viral infections, certain types of cancer and autoimmune disorders,[119] as well as skin disorders, as cutaneous cannabinoid (“c[ut]annabinoid”) signaling is deeply involved in the maintenance of skin homeostasis, barrier formation and regeneration, and its dysregulation is implicated to contribute to several diseases and disorders, e.g., atopic dermatitis, psoriasis, scleroderma, acne, hair growth and pigmentation disorders, keratin diseases, various tumors, and itch, as the endocannabinoids (eCBs; e.g., NAE 20:4; AEA), the eCB-responsive receptors (e.g., CB1, CB2), as well as the complex enzyme and transporter apparatus involved in the metabolism of the ligands, show to be expressed in several tissues, including the skin,[120] where 18 prostanoids, 12 hydroxy-fatty acids, 9 endocannabinoids and N-acyl ethanolamides (NAEs), and 21 non-hydroxylated ceramides and sphingoid bases, of which several demonstrating significantly different expression in the tissues assayed, demonstrate the diversity of lipid mediators involved in maintaining tissue homeostasis in resting skin and hint at their contribution to signaling, cross-support, and functions of different skin compartments.[121]
The NAE substitutes, the phytocannabinoids from the flowers and fruits, like the psychoactive compound Δ9-tetrahydrocannabinol (THC: C21H30O2) and the nonpsychotropic compounds cannabidiol (CBD: C21H30O2), and leaves (THCA/CBDA: C22H30O4),[122] from the plant, are also potent PPARγ agonist with neuroprotective activity,[123][124][125] and found to modulate inflammatory responses by regulating the production of cytokines from keratinocytes in several experimental models of skin inflammation, by CB2 and TRPV1 activation, where CBD dose-dependently elevates the levels of NAE 20:4 (AEA) and inhibits poly-(I:C)-induced release of MCP-2, interleukin-6 (IL-6), IL-8, and tumor necrosis factor-α, in allergic contact dermatitis (ACD),[126] through the endocannabinoid system (ECS), and where FAAH–deficient mice, which have increased levels of NAE 20:4, displayed reduced allergic responses in the skin,[127] as the activation of CB1 or CB2 increases endocannabinoid levels by inhibiting fatty acid amide hydrolase (FAAH) or adenylyl cyclase, and activation of CB1 is tightly associated with the generation of cellular ceramides.[13]
Beside the CB1 receptors being triggers of the generation of ceramides that mediate neuronal cell fate, the skin CB1 receptor aktivation also increases ceramides, with long-chain fatty acids (FAs) (C22–C24), which mainly account for the formation of the epidermal barrier, through activation of ceramide synthase, CerS 2 and CerS 3, thereby resulting in the enhancement of epidermal permeability barrier function in IL-4 inflamed skin.[13]
Synthesis of LC-PUFAs in humans and many other eukaryotes starts with:
* Linoleic acid (LA: C18H32O2; 18:2-n6) → Δ6-desaturation (removing two hydrogen atoms, creating a double bond and a bend in the fatty acid) → γ-linolenic acid (GLA: C18H30O2; 18:3-n6) → Δ6-specific elongase (introducing two carbons and four hydrogens) → Dihomo-gamma-linolenic acid DGLA: C20H34O2; 20:3-n6) → Δ5-desaturase → arachidonic acid (AA: C20H32O2; 20:4-n6) → also endocannabinoids.
* α-Linolenic acid (ALA: C18H30O2; 18:3-n3) → Δ6-desaturation → stearidonic acid (SDA: C18H28O2; 18:4-n3) and/or → Δ6-specific elongase → eicosatetraenoic acid (ETA: C20H32O2; 20:4-n3) → Δ5-desaturase → eicosapentaenoic acid (EPA: C20H30O2; 20:5-n3).
By a Δ17-desaturase, gamma-Linolenic acid (GLA: C18H30O2; 18:3-n6) can be further converted to stearidonic acid (SDA: C18H28O2; 18:4-n3), dihomo-gamma-linolenic acid (DHGLA/DGLA: C20H34O2; 20:3-n6) to eicosatetraenoic acid (ETA: C20H32O2; 20:4-n3; omega-3 Arachidonic acid)[128] and arachidonic acid (AA: C20H32O2; 20:4-n6) to eicosapentaenoic acid (EPA: C20H30O2; 20:5-n3), respectively.[129]
- Fatty acids with at least 20 carbons (C20) and three double bonds (20:3) bind to CB1 receptors.[1]
- Arachidonic acid (AA) is also the catalyst to the formation of the two main endocannabinoids, Anandamide (AEA) and 2-arachidonoylglycerol (2-AG).
* Anandamide (AEA: C22H37NO2; 20:4,n-6) is an N-acylethanolamine resulting from the formal condensation of the carboxy group of arachidonic acid (AA: C20H32O2; 20:4-n6) with the amino group of ethanolamine (C2H7NO), bind preferably to CB1 receptors.[130]
* 2-arachidonoylglycerol (2-AG: C23H38O4; 20:4-n6) is an endogenous agonist of the cannabinoid receptors (CB1 and CB2), and the physiological ligand for the cannabinoid CB2 receptor.[131] It is an ester formed from omega-6-arachidonic acid (AA: C20H32O2; 20:4-n6) and glycerol (C3H8O3),[132] and tissue levels of 2-AG is usually several tens to several hundreds of times those of AEA.[51]
The N-acylethanolamine synthesis pathway is one of the non-oxidative pathways in which the mainly fish oil derived n−3 long-chain polyunsaturated fatty acids (n−3 LCPUFA, C18 to C22), ω-3 FAs docosahexaenoic acid (DHA: C22H32O2; 22:6, n-3) and eicosapentaenoic acid (EPA: C20H30O2; 20:5, n-3) are converted to NAE 22:6 docosahexaenoyl ethanolamide (DHEA: C24H37NO2; 22:6, ω-3)[38] and NAE 20:5 eicosapentaenoyl ethanolamide (EPEA: C22H35NO2; 20:5, ω-3), or Anandamid (20:5, n-3),[44] by incorporated ethanolamine (MEA: C2H7NO), respectively. On common diseases including cancers, this conversion show beneficial synergistic effect, when administered with the NAE-fatty acid familiar cannabinoids,[40] like delta(9)-tetrahydrocannabinol (THC: C21H30O2), - a metabolite of delta9-tetrahydrocannabinolic acid (THCA: C22H30O4), a diterpenoid, with a carboxyl group (–COOH) at one end, like 11-nor-9-carboxy-THC (THC-COOH: C21H28O4), the secondary metabolite of THC, which is formed in the body after cannabis is consumed, that has a role as an anti-inflammatory and a neuroprotective agent,[133] - and a non-narcotic analgesic, a hallucinogen, a cannabinoid receptor agonist and an epitope.[134] As Cannabis sativa and their derivatives act in the organism by mimicking endogenous substances, the endocannabinoids, that activate specific cannabinoid receptors, is why cannabinoids are found to be selective antitumour compounds, that can kill tumour cells, by growth arrest or apoptosis, without affecting their non-transformed counterparts, is probably because, cannabinoid receptors regulate cell-survival and cell-death pathways differently in tumour and non-tumour cells.[135][136][137][138][139]
Metabolic production of NAEs
Diets in mammals, containing 20:4,n−6 and 22:6,n−3, are found to increase several biologically active NAEs in brain homogenates as metabolic products, like 20:4,n−6 NAE (4-fold), 20:5,n−3 NAE (5-fold), and 22:5,n−3 and 22:6,n−3 NAE (9- to 10-fold). The increase in all of the metabolic NAEs is regarded biologically important, because NAEs having fatty acids with at least 20 carbons and three double bonds bind to CB1 receptors,[1] and endogenously released NAE 20:4 and 2-arachidonylglycerol (2-AG: C23H38O4; 20:4,n-6), the ester metabolic formed from omega-6-arachidonic acid (AA: C20H32O2; 20:4, n-6) and glycerol (C3H8O3), are also found to activate CB2 receptors in addition,[34] where 2-AG is the physiological ligand.[131]
The hydrolysis of NAE to free fatty acid (FFA) and ethanolamine (MEA) in animals, is catalyzed by fatty acid amide hydrolase (FAAH) or by a N-acylethanolamine-hydrolyzing acid amidase (NAAA), and the polyunsaturated NAEs such as NAE 18:2, NAE 18:3, or NAE 20:4 can also be oxygenated via lipoxygenase (LOX) or cyclooxygenase (COX), to produce ethanolamide oxylipins, like prostaglandin ethanolamides (prostamide) by COX-2, with various potential bioactivities that may have enhanced affinity with cannabinoid receptors in comparison to their respective non-oxygenated NAEs,[140][54][141] as well as to oxygenated eicosanoid ethanolamides, prostaglandins, and leukotrienes, all believed to be important signaling compounds.[142]
The major COX-2 derived prostanoid product from NAE 20:4 (AEA) are prostaglandin E2 (PGE2) ethanolamide (PGE2-EA; prostamide E2) and PGD2 ethanolamide (PGD2-EA; prostamide D2), might have many important functions,[143] as PGE2 and PGD2 are pro-inflammatory mediators responsible for the induction of inflammation,[140] PGE2-EA and PGD2-EA are contrary both growth inhibitory and can induce apoptosis,[144] as well as that NAE 20:4 (AEA) and/or its prostamide metabolites in the renal medulla, may represent medullipin and function as a regulator of body fluid and the mean arterial pressure (MAP).[145]
Fatty acid amide hydrolase (FAAH) is the main degrading enzyme of NAE 20:4 (AEA) and NAE 18:1 (OEA), which have opposite effects on food intake and energy balance. AEA, an endogenous ligand of CB1 cannabinoid receptors, enhances food intake and energy storage, whereas OEA binds to peroxisome proliferator-activated receptors-α to reduce food intake and promoting lipolysis, thereby FAAH deficiency promotes energy storage and enhances the motivation for food, through the enhancement of AEA levels rather than promoting the anorexic effects of OEA.[146] Tetrahydrocannabinol (THC: C21H30O2) is found to lower production of NAE 20:4 (AEA) and 2-AG, that is synthesized in an on-demand manner when needed for activation,[147] by a biphasic response after THC injection reaching maximal values at 30 min., where AEA increased slightly from 0.58 ± 0.21 ng/ml at baseline to 0.64 ± 0.24 ng/ml, and 2-AG from 7,60 ± 4,30 ng/ml to 9,50 ± 5,90 ng/ml, and after reaching maximal concentrations, EC plasma levels decreased markedly to a nadir of 300 min after THC administration to 0.32 ± 0.15 ng/ml for AEA, and 5,50 ± 3,01 ng/ml for 2-AG, and returned to near baseline levels until 48 hours after the experiment, in 25 healthy volunteers who received a large intravenous dose of THC (0.10 mg/kg).[74]
Insulin medication and intraoperative doses of insulin is also found, but not recognized by companies producing and selling medication to general public also as a slimming formula like Wegovy, to get its anorectic effect by the involvement of FAAH activity, which, beside of other NAE's, degrade NAE 20:4 (AEA),[148] suggest that insulin may play a key role in the obesity-linked dysregulation of the adipose ECS at the gene level.[149] And is possible why the European Medicines Agency (EMA) in 2023 are investigating several reports from European countries about suicidal thoughts and thoughts of self-harm in patients, who have been treated with Novo Nordisk's popular medicines for obesity and diabetes.[150] An outcome also seen in the CB1 receptor blocker rimonabant, an anorectic antiobesity drug that was first approved in Europe in 2006 but was withdrawn worldwide in 2008 due to serious psychiatric side effects,[122][86] and happening at the same time as EMA, has raised a safety alert for Wegovy, that also applies to the companies diabetes medication Ozempic, based on a study that suggests that the active substance in the two preparations, can increase the risk of thyroid cancer in patients with type 2 diabetes.[151]
FAAH expression, that metabolizes NAE 20:4 (AEA) involved in the regulation of emotional reactivity, into ethanolamine and arachidonic acid, is found significantly increased in depressive-like phenotypes, where knockout or pharmacological inhibition of FAAH effectively reduces depressive-like behavior, with a dose-dependent effect, that elicits anxiolytic and antidepressant-like effects, like the NAE 20:4 (AEA) substitutes ∆9-THC and other cannabinoids that may contribute to the overall mood-elevating properties of cannabis,[95][152][153] and differences in FAAH expression in depressive-like phenotypes were largely localized to animal prefrontal cortex (PFC), hippocampus and striatum, containing high densities of CB1 receptors.[154][155] As well as FAAH levels in amygdala and PFC are elevated in borderline personality disorder, which relates to the hostility and aggression, are consistent with the model, that lower endocannabinoid tone perturb PFC circuitry that regulates emotion and aggression, provide preliminary evidence of elevated PFC FAAH binding in any psychiatric condition.[88]
A FAAH 385A mutant alleles have been found to have a direct effect on elevated plasma levels of NAE 20:4 (AEA) and related NAEs in humans, and biomarkers that may indicate risk for severe obesity that suggest novel ECS obesity treatment strategies,[156] as leptin increases the FAAH activity and reduces NAE 20:4 (AEA) signaling, particularly within the hypothalamus, to promote a suppression of food intake, a mechanism that is lost in diet-induced obesity and modulated by a human genetic variant (C385A) of the FAAH gene.[157] The cannabinoid type 1 receptors (CB1) and their endogenous ligands, the endocannabinoids, present in peripheral organs, such as liver, white adipose tissue, muscle, and pancreas, where it regulate lipid and glucose homeostasis, and dysregulation of it, has been associated with the development of obesity, characterized by chronic mild inflammation,[158] and its sequelae, such as dyslipidemia and diabetes, are involved in modulating food intake and the motivation to consume palatable food.[159]
NAE 20:4 related THC treatment have shown to increase culture protein content and reduced methyl-(3)H-thymidine incorporation, and cells treated with THC underwent adipogenesis shown by the expression of PPARγ and had increased lipid accumulation. Basal and IP-stimulated lipolyses were also inhibited by THC, and the effects on methyl-(3)H-thymidine incorporation and lipolysis seem to be mediated through CB1- and CB2-dependent pathways. THC did also decrease NAPE-PLD, the enzyme that catalyzes and converts ordinary lipids into chemical signals like NAE 20:4 (AEA) and NAE 18:1 (OEA), in preadipocytes and increased adiponectin and TGFβ transcription in adipocytes, results that show the ECS interferes with adipocyte biology and may contribute to adipose tissue (AT) remodeling. And this stimulation of adiponectin production and inhibition of lipolysis from THC may be in favor of improved insulin sensitivity under cannabinoid influence.[160]
A full agonist at the CB1 receptor is found able to up-regulate PPARy, and increased (+50%) glucose uptake, the translocation of glucose transporter 4, and intracellular calcium in fat cells, that indicate a role for the local endocannabinoids in the regulation of glucose metabolism in human adipocytes and suggest a role in channelling excess energy fuels to adipose tissue in obese humans.[161] This is consistent with the decreased prevalence of diabetes seen in marijuana users,[162] and significantly reduced body mass index (BMI) and rates of obesity in Cannabis users,[163][164] as endocannabinoids modulate pancreatic β-cells function, proliferation, and survival, as well as insulin production, secretion, and resistance, where animal and human research suggest that increased activity of the endocannabinoid system, may lead to insulin resistance, glucose intolerance and obesity.[165]
Consistent with the associated reduced prevalence of non-alcoholic fatty liver disease (NAFLD) among cannabis users, that find significantly lower NAFLD prevalence compared to non-users, i.e. 15% lower in non-dependent users and 52% lower in dependent users, and dependent patients had 43% significantly lower prevalence of NAFLD compared to non-dependent patients.[166] And also by using multivariable logistic regression, and after adjusting for potential confounders, patients with cannabis abuse (daily consumption) is found 55% less likely to have hepatocellular carcinoma (adjusted odds ratio, 0.45, 95% confidence interval, 0.42-0.49) compared with patients without cannabis abuse in the periode 2002 - 2014.[138]
In addition to metabolism by FAAH, COX-2 and LOXs, NAE 20:4 (AEA) can also undergo oxidation by several of human cytochrome P450 (CYPs) enzymes, resulting in various oxidized lipid species, some of which have biological relevance as CYP-derived epoxides, that can act as a potent agonist of CB2 receptors.[142]
NAE 20:4 (AEA: C22H37NO2) which is similar in structure to N-arachidonoyl glycine (Nagly: C22H35NO3 - a carboxylic acid COOH) are metabolically interconnected, as oxidation of the hydroxyl group of NAE 20:4 (AEA) leads to NAgly, preferring G-protein coupled receptor (GPR) 118, with a molecular structure, that are found of pharmacological interest, as region one confers a high degree of specificity of action, as polyunsaturated residues produce molecules with analgesic and anti-inflammatory action, of which saturated structures, are inactive. Region two is related to metabolic stability as NAgly is degraded by FAAH activity. And last, region 3, the amino acid residue, can have an effect on the analgesic and anti-inflammatory activities depending on steric factors and the chiral nature of the amino acid.[167] Also the amino acid residue at 296 and the hydroxyl groups of THC, 11-hydroxy-THC (11-OH-THC: C21H30O3) are critical for potentiation of glycine receptors (GlyRs) and for some of the cannabis-induced analgesic and therapeutic effects.[75]
It is also found that long-chain fatty acid conjugates from the metabolic hydroxyl oxidation product of the phytocannabinoid, THC (C21H30O2), 11-hydroxy-THC (11-OH-THC: C21H30O3) are proposed to be a form in which THC may be stored within tissues.[168][169] And the last cytochrome P450 oxidation product of THC afford the non-psychoactive and long-living 11-nor-9-carboxy-THC (THC-COOH: C21H28O4) as main metabolite, that in some authors’ opinion, are insufficiently characterized, as an acid metabolite seen as a final product in both cannabis-plants and mammals, with their main unanswered questions, "Could any of the pharmacological effects observed for THC be attributed to THCA (C22H30O4) and/or THC-COOH, and could THC also be a potential pro-drug to another pharmacological entity?".[170][167][171][172][173]
NSAIDs that inhibit COX2, may find its medical influence from the cannabinoid system, either by inhibiting the breakdown of NAE 20:4 (AEA) by FAAH (i.e. ibuprofen, indomethacin, flurbiprofen, ibu–am5), or by inhibiting a possible intracellular transporter of endocannabinoids (i.e. acetaminophen).[174]
The phytocannabinoid THC is found to have twenty times the anti-inflammatory potency of aspirin and twice that of hydrocortisone, but in contrast to NSAIDs, it demonstrates no COX inhibition at physiological concentrations.[175]
Another of the main phytocannabinoids, cannabidiol (CBD: C21H30O2) is found to produce a significant increase in serum NAE 20:4 (AEA) levels, by inhibiting the intracellular degradation catalyzed by FAAH, suggest the inhibition of NAE 20:4 (AEA) deactivation may contribute to the antipsychotic effects of CBD, potentially representing a mechanism in the treatment of schizophrenia, with a markedly superior side-effect profile, compare to amisulpride, a potent antipsychotic. CBD were also seen to elevate serum levels of the non-cannabimimetic lipid mediators, NAE 16:0 (PEA) and NAE 18:1 (OEA), but amisulpride did not.[176][177]
FAAH inhibitors are seen to both increase alcohol consumption (NAE 20:4; AEA) and prevent against oxidative stress caused by binge ethanol consumption, and as NAE 16:0 (PEA) and NAE 18:1 (OEA), through the endocannabinoidome-related peroxisome proliferator-activated receptor-α (PPAR-α) is involved in the actions of NAEs with no endocannabinoid activity, have been reported to exhibit neuroprotective effects, suggest a strengthening of the ECS may reflect a homeostatic mechanism to prevent the neurotoxic effects induced by alcohol with a relevant role of other non-cannabinoid congeners in the alcohol exposure, and the further activation in response to the negative affective state, like the anxiety,[178] associated to alcohol withdrawal.[179] Or poorer recall of verbal and nonverbal information, as well as reduced visuospatial skills related to alcohol hangover and withdrawal symptoms in youth, a relationship not seen in adolescents with similar levels of alcohol involvement if they are heavy users of marijuana.[180]
The cannabinoid CB1 receptor play a critical role in mediating the adolescent behavior, because enhanced CB1 density and endocannabinoid (eCB) signaling occur transiently during the periode from childhood to adolescence and reverse when adult and mature in normal phenotypes. Reports on enhanced adolescent CB1 signaling, suggest a pivotal role for the CB1 in an adolescent brain as an important molecular mediator of adolescent behavior, as adult CB1 mutant rats exhibit an adolescent-like phenotype with typical high risk seeking, impulsivity, and augmented drug and nondrug reward sensitivity, by an instinctive need or call for activation, and partial inhibition of CB1 activity normalized behavior and led to an adult phenotype, is why it is concluded that the activity state and functionality of the CB1 is critical for mediating adolescent behavior and further turn to an adult phenotype, by normal CB1 downscaling.[181] This is also do to the cannabinoid system and its neurotransmitter NAE 20:4 (AEA), that highly participate in the modulation of human states and appropriate human emotional responses by activation of the CB1 receptor,[178][182] also found in frontal neocortical areas, subserving higher cognitive and executive functions, and in the posterior cingulate, a region pivotal for consciousness and higher cognitive processing.[183][184][185]
Acute administration of ethanol inhibits receptor-mediated release of NAE 20:4 (AEA), whereas chronic ethanol administration increases levels of AEA that participates in the neuroadaptations associated with chronic ethanol exposure, as the inhibition of AEA release by acute ethanol administration, not derive from increased fatty acid ethanolamide degradation by FAAH.[186][187]
However, alcohol (EtOH) is seen to increase levels of NAE 20:4 (AEA), and its precursor N‐arachidonoylphosphatidylethanolamine (N‐ArPE), a glycero-phospho-ethanolamine,[188] significantly, that may be a mechanism for neuronal adaptation and serve as a compensatory mechanism to counteract a continuous presence of EtOH, that together with previous results indicate the involvement of the endocannabinoid system in mediating some of the pharmacological actions alleged of EtOH, also seen in red wine components,[189] and in Humulus lupulus to preserve and flavor beer, widely cultivated for use by the brewing industry, through caryophyllene, a dietary cannabinoid,[190] that is a selective full agonist at CB2 and also act through PPAR nuclear receptors (i.e. PPARα and PPARγ), with countless beneficial and non-psychoactive effects,[191][192][193] that may constitute part of a common brain pathway mediating reinforcement of drugs of abuse including EtOH,[194] by elevated CB1.[195] The CB1 receptor binding is 20-30% lower in patients with alcohol dependence than in control subjects in all brain regions and is negatively correlated with years of alcohol abuse, and the CB1 receptor binding remain similarly reduced after 2–4 weeks of abstinence, suggests an involvement of CB1 receptors in alcohol dependence in humans.[196]
Similar pathways of hydrolysis or oxidation of NAEs are also found in plant cells.[197][198]
NAE system in plants
N-acylethanolamines (NAEs), constitute a class of lipid compounds naturally present in both animal and plant membranes, as constituents of the membrane-bound phospholipid, N-acylphosphatidylethanolamine (NAPE). NAPE is composed of a third fatty acid moiety linked to the amino head group of the commonly occurring membrane phospholipid, phosphatidylethanolamine.[34]
A study in 2000 find, that higher plants use defense signaling, to combat cellular stressful situations (homeostasis), like in osmotic stress, where high levels of NAEs after a periode of dehydration, are metabolized fast during the first few hours of imbibition, and in response to pathogen elicitors, that lead to signal transduction and membrane protection, in the same way as several mammalian cell types, coupled to endocannabinoid signaling, do, by releasing saturated and unsaturated long-chain NAEs, and saturated medium-chain NAEs, that can act as lipid mediators to modulate ion flux and activate defense gene expression.[199]
The levels of NAEs increases 10- to 50-fold in tobacco (Nicotiana tabacum) leaves treated with fungal elicitors, as a protection against it, by producing the N-myristoylethanolamine (Myristamide-MEA: C16H33NO2; NAE 14:0), that specific binds to a protein in tobacco membranes with biochemical properties appropriate for the physiological responses, and it do not show identical binding properties to NAE-binding proteins in intact tobacco microsomes, compared to non-intact microsomes. In addition to this, antagonists of mammalian CB receptors was seen to block both of the biological activities previously attributed to NAE 14:0, this endogenous NAE that is accumulated in tobacco cell suspensions and leaves after pathogen elicitor perception, is why it is proposed, that plants possess an NAE-signaling pathway with functional similarities to the “endocannabinoid” pathway of animal systems, and this pathway, in part, participates in xylanase elicitor perception in the tobacco plant, as well as in the Arabidopsis and Medicago truncatula plant tissues.[34]
Medical values
N-acylethanolamines (NAEs), with its cell-protective and stress-combating action-response of organisms, also produced in neurons, together with N-acyl-phosphatidylethanolamine (NAPE), in response to the high intracellular Ca2+ concentrations that occur in injured neurons,[200] have shown promise as therapeutic potential in treating bacterial, fungal, and viral infections, as NAEs also exhibit anti-inflammatory, antibacterial, and antiviral properties, which have considerable application potential.[22]
In pediatric medicine for conditions including “non-organic failure-to-thrive” and cystic fibrosis.[67] A dysfunction of the endocannabinoid system is researched for a possible determining factor for causing infertility in cystic fibrosis (CF), as the illness is associated with an imbalance of fatty acids, show that mild stimulation of the endocannabinoid system (CB1 and CB2) in infancy and adolescence, appears to normalize many reproductive processes and prevent infertility in CF males. The mild stimulated, were fully fertile, producing offspring comparable by the number of litters and the number of pups as the wild-type mice, and their counterparts, not treated, were shown completely infertile.[201]
As NAE related Cannabis has an ancient tradition of usage as a medicine in obstetrics and gynecology, its extracts, may represent an efficacious and safe alternative for treatment of a wide range of conditions in women including dysmenorrhea, dysuria, hyperemesis gravidarum, and menopausal symptoms.[202]
It has been found that social contact increases, whereas isolation decreases, the production of the endogenous marijuanna-like neurotransmitter, NAE 20:4 (AEA), in nucleus accumbens (NAc), which regulate motivated behavior, and this NAE 20:4 (AEA) production is via oxytocin, the neuropeptide reinforcing parental and social bonding. Activation of CB1 cannabinoid receptors in NAc, are necessary and sufficient to express the rewarding properties of social interactions, i.e. social contact reward.[89][203][204] In addition, CB1 activation also suppresses release of serotonin, dopamine, acetylcholine and noradrenaline, which are mediating the characteristic cognitive and antidepressant effects.[205] As well as norepinephrine release,[206] as it is suggested that a major function of the ECS also lies in buffering the symphatico-adrenergic response to stress.[207]
To use in expected global heating scenario, in a catastrophic “hothouse Earth,” possible well beyond the control of humans,[208][209][210] where "wet bulb temperatures," taken by a thermometer wrapped in a wet cloth, show temperatures of 35C or higher, and considered the limit to human survival and heighten humidity makes it harder for people to cool down via sweating,[211][61] coursed by the pollution of the troposphere, that tight holds 99% of human made solid particle pollution, and keeps CO2 in it for more than 100 years,[212][213][214] for citizens who can't afford an air-condition unit, to cool down and prevent heatstroke with an elevated core body temperature above 40 °C with neurologic dysfunctions, that can lead to a syndrome of multiple organ defect,[215] and cell stress, as it is found, that the CB1 receptor activation, here by a phytocannabinoid Δ9-THC administration, induces profound hypothermia, that is rapid in onset, persistent for 3–4 hours, dose-dependent and is accompanied by a reduction in oxygen (O) consumption, which indicate reduced heat production, as opposed to increased heat loss.[216][217][218]
* → THCA:COOH: C22H30O4 (heating/storage) → THC: C21H30O2 → THC-OH: C21H30O3 → THC:COOH: C21H28O4 → profound hypothermia, a lowering of body temperature,[219] accompanied by a reduction in oxygen consumption.
In metabolism of THCA from fresh plant material used orally, is conversion to Delta9-THC not observed:[220]
THCA:COOH: C22H30O4 → THC-OH: C21H30O3 → THC:COOH: C21H28O4
To be protected where head injury is a possibility, as a positive THC screen is associated with significant decreased mortality in adult patients sustaining traumatic brain injury (TBI), as research work, by a 3-year retrospective review of registry data at a Level I center of patients sustaining TBI, find mortality in the THC(+) group (2.4% [2 patients]) significantly decreased compared with the THC(-) group (11.5% [42]) in 446 cases meeting all inclusion criteria.[172] And further have shorter hospital length of stay (LOS) and shorter ventilator days, than THC(-) patients sustaining TBI. For severely injured trauma patients with Injury Severity Score ≥16, a THC(+) screen show significantly lower intensive care unit LOS and mortality (19.3% versus 25.0%) than THC(-) patients, shown by 4849 patients included at two large regional trauma centers between 2014 and 2018.[221]
As the fatty acid amide hydrolase (FAAH) have showed significant decreased in bhang users as compared to controls, and indicating that the decrease in FAAH protein level is closely related to the duration of bhang use, and further revealed that the bhang–induced immunotoxicity, could be attributed to decrease in FAAH protein, bhang could also be a healthy drink/preparation to suppress an overactive immune response.[173][222]
Fatty acid amide hydrolase (FAAH) inhibition has been found neuroprotective with therapeutic potential against neuropathological states including traumatic brain injury, Alzheimer's, Huntington's, and Parkinson's diseases, and stroke.[223]
A molecular mechanism through which NAE 20:4 (AEA) plant competitive substitute THC cannabinoid molecules can affect the development of Alzheimer's disease, the leading cause of dementia,[224] or its impact:
THC: C21H30O2 → THC-OH: C21H30O3 → THC:COOH: C21H28O4 → a significantly superior inhibitor of Amyloid beta (Aβ) aggregation and tau phosphorylation, compared to approved drugs prescribed for the treatment of Alzheimer's disease in 2008, through which these molecules directly can affect the development by activation of both CB1 and CB2 receptors, which inhibit the enzyme acetylcholinesterase (AChE), which further prevent AChE-induced amyloid β-peptide (Aβ) aggregation, as they also are able to bind to the anionic site of AChE, a region involved in and critical for amyloid formation, as well as by promoting the brain's intrinsic repair mechanisms, and promote neurogenesis, endocannabinoid signaling has demonstrated to modulate numerous concomitant pathological processes, including neuroinflammation, excitotoxicity, mitochondrial dysfunction, and oxidative stress.[225][226][227] However other phytochemicals that are present in Cannabis sativa is found to interact with each other in a synergistic fashion, called the entourage effect, that seems to have greater therapeutic potential when administered together, rather than individually.[228][229][134][230]
A synergistic outcome that also show different cannabinoids can be effective against harmful bacteria including those that are resistant to common antibiotics, like Methicillin-resistant Staphylococcus aureus (MRSA) causing various types of life-threatening infections, such as septic shock, endocarditis and severe pneumonia, coursed by the misuse of antibiotics, which is the leading cause of the emergence of antibiotic-resistant bacteria. They do so by inhibit the formation of biofilms and also eradicate pre-existing ones, was showcased in 1976, where it was discovered that THC and CBD can be used as bacteriostatic agents and are able to kill a panel of human pathogenic strains, and later a panel of cannabinoids are found able to do the same in different bacteria strains.[231][232]
Different medication and intervention regimes, and lifestyle modifications, like diet, weight control, exercise, mindfulness as yoga and meditation, and the use of psychoactive substances, like alcohol,[194] tobacco, coffee,[233][234] and cannabis, beside general anaesthesia regimens (i.e. propofol, etomidate, sevoflurane, isoflurane, sufentanil),[235][236] and Insulin medication and intraoperative doses of insulin,[148][149] etc, do also modulate it, either by being a FAAH inhibitor, that blocks the breakdown of NAE 20:4 (AEA), and/or enhance or lowering its production, and/or by activate or inactivate the receptors connected, as arachidonic acid (C20H32O2; 20:4, ω-6), the precursor of NAE 20:4 (AEA) and other eCBs, is present in every cell membrane of the body, and their on demand synthesis is regulated by electrical activity and calcium (Ca2+) shifts.[52][237][238][239][240][241][172][242]
The evolutionary harm reduction program
The central thesis is, that harm reduction is not only a social concept,[243] but also a biological one. More specifically, evolution does not make moral distinctions in the selection process, but utilizes a cannabis-based approach, seen from the oldest pollen, where Cannabis and Humulus diverged between 18.23 mya and 27.8 mya ago, and consistent with Cannabis dated to 19.6 ago (Ma), in northwestern China, and converge on the northeastern Tibetan Plateau, in the general vicinity of Qinghai Lake, which is deduced as the Cannabis centre of origin, and co-localizes with the first steppe community that evolved in Asia,[244] or Yunnan, in the southwest of China, also identified as "the birthplace of tea ... the first area where humans figured out that eating tea leaves or brewing a cup could be pleasant", and helpful, by its cannabimimetic bioactivity of catechin derivatives occurring in tea leaves, as the region of origin,[245][246] to harm reduction in order to promote survival of the fittest. Evidence provided from peer-reviewed scientific literature supports the hypothesis, that humans, and all animals,[247] since the primordial CB receptor evolved at least 600 million years ago; a date that broadly consistent with the Cambrian explosion,[248] make and use internally produced cannabis-like products (endocannabinoids) as part of the evolutionary harm reduction program. More specifically, endocannabinoids homeostatically regulate all body systems (cardiovascular, digestive, endocrine, excretory, immune, nervous, musculo-skeletal, reproductive),[237] as a versatile tool available to organisms to fine-tune homeostasis,[249] and modulating endocannabinoid activity have therapeutic potential in almost all diseases affecting humans.[122][250][251][252][253] Therefore, the health of each individual is dependent on this system are working appropriately,[64][102] and imagine what could be achieved if signaling through these receptors could be controlled: happy, slim, and healthy people who remember that they're pain-free,[254][206] by forgetting,[255][256] and ignore it,[257] achieved though cannabis, the evolutionary byproduct of a plant that evolved as a systemic homeostasis to affect the ECS and to become its natural key, stemming back to aquatic species 400 million years before the arrival of plants and trees.[229][122]
Cosmic birth of NAE elements by star death
NAE 20:4 (AEA: C22H37NO2; 20:4, ω-6) is the amide of arachidonic acid (C20H32O2; 20:4, ω-6) and ethanolamine (MEA: C2H7NO).
NAE 20:5 (EPEA: C22H35NO2; 20:5, ω-3) or Anandamide (20:5, n-3). It is the amide of eicosapentaenoic acid (EPA: C20H30O2; 20:5, ω-3)
NAE 22:6 (DHEA: C24H37NO2; 22:6, ω-3), or Anandamide (22:6, n-3) "synaptamide", is the amide of docosahexaenoic acid (DHA: C22H32O2; 22:6, ω-3) and ethanolamine (MEA: C2H7NO)
In the heart of a heavy star, like Betelgeuse, and in its core, is where all the ingredients of life (and elements of NAEs) are found made. Deep in its core, the star will fight a futile battle against its own gravity, as it tries to stop itself collapsing under its own weight, is where new elements are made in a sequence of separate stages.
Stage one, is while there is still a supply of hydrogen (H) to burn. As the star burn in hydrogen to helium (He) in the core, are vast amounts of energy released and that energy escapes and thereby creating an outward pressure, which balances the force of gravity, and holds the star off and keeps it stable. But the hydrogen (H) in the core will eventually run out, and the fusion reactions will stop and no energy will be released, and the outward pressure will disappear, and the core will start to collapse and very rapidly leaving a shell of hydrogen (H) and helium (He) behind.
Beneath this shell, as the core collapses, the temperature rises again until at a hundred million degrees, the stage two stars and helium nuclear begin to fuse together. Helium fusion does two things. First, it releases more energy so the collapse is halted, and secondly it produce two more elements in that process. Carbon (C) and oxygen (O), two elements vital for life (and molecules in the NAE structure), with further collapses until it eventually form iron (Fe). And when that happens, the star collapses around itself. And, through a nebula, meteorites and comet impact, this is where all the carbon, hydrogen, oxygen etc. in the NAE molecular structure come from, and every atom particle in every living thing on Earth, was probably produced in and carried from the elements of a dying star.[258]
And due to the chemical ability to build chains and rings, carbon (C) forms with quite a few other elements, like hydrogen (H), oxygen (O), nitrogen (N), phosphorus (P) and sulfur (S) the basis of the myriad of the chemical compounds, proteins, DNA, carbohydrates, lipids, etc. of living organisms.
NASA-funded scientists have evidenced, that some building blocks of DNA, a nucleic acid, and the molecule that carry genetic instructions for life, found in meteorites, were likely created in space. The research supports the theory, that a "kit" of finished parts created in space and delivered to Earth by meteorite and comet impact, has aided the origin of life.[259]
A simulated Comet impact on the surface of the Earth, as it probably would look like 3.8 billion years ago (3800 MYA) at the late heavy bombardment, show that amino acids (i.e. organic compounds arisen from amine (-NH2) and carboxylic acid (-COOH) functional groups, producing proteins) in water is able to make peptide bonds (i.e. bonds from the carboxyl group of one amino acid to the amino group of the other amino acid), and peptides from the heat of the impact through Earth's atmosphere, and this could be a spark to early life on Earth.[260]
Using the samples returned by the spacecraft Stardust, other scientists discovered the amino acid glycine (C2H5NO2) fully formed in a comet tail, but this glycine was not like this on Earth; it was built with some subtle differences - traces of COOH's important presence and interstellar heritage, as well as strengthened the theory of panspermia, which claims that these "seeds" of life are widespread throughout the universe,[261] and among the most widespread and versatile signaling molecules ever discovered.[262]
Further other scientists have used the radio telescope (GBT) in West Virginia to study a giant cloud of gas about 25,000 light-years from Earth, near the center of the Milky Way. The chemicals found in the cloud include one molecule that is thought to be a precursor to a key element in DNA and another that may have a role in the formation of the amino acid, alanine (Ala: C3H7NO2).
Cyanomethanimine (C2H2N2), is a step in the process that is believed to produce adenine (C5H5N5), one of the four nucleobases that make up the "step" of the ladder-like structure of DNA. The second molecule, called ethanamine (C5H5N), is thought to play a role in the formation of alanine (Ala - C3H7NO2), one of the twenty amino acids in the genetic code.[263][264]
* → An atom consists of a nucleus, the atom nucleus, and a shell. The atom's positively charged particles, protons, lie in the nucleus and consist of the smallest building blocks we know of in the universe, called quarks, which are held together by gluons, are everywhere. Along with the protons are neutrons, which are neutral. Outside the nucleus is the mantle, where the atom's negatively charged particles, electrons, interact (in the form of standing waves), and is the absolute smallest part of an element, that can exist independently, but is most often bound to other atoms that can either belong to the same element or other elements, thereby forming a molecule.[265]
With a spike on a graph, an entourage effect and retrograde signaling, by the motto “Science for Peace", and, also in the spirit of Niels Bohr, the Danish physicist with the first relatively correct model of how an atom looks like, - an international group of about 6000 researchers connected at CERN, by a particle accelerator located 100 meters underground, called LHC Large Hadron Collider 1, which consists of a 27 km long tube in a circle, that can accelerate protons to near the speed of light and smash them together, demonstrated on 4 July 2012 the existence of the Higgs particle, a manifestation of the existence of the Higgs field - an invisible field in the entire Universe (symmetry breaking), believed to have emerge at about 1 picosecond (10−12 s) after the Big Bang, and required for atoms and other structures to form, as well as for nuclear reactions in stars, such as the Sun and Betelgeuse. As so, the Higgs field is responsible for this symmetry breaking, and which all particles pass through, and which provide them their mass and thereby, weight, and without this field, all particles and elements in the NAE-structure, would be moving around at the speed of light, because they didn't have a mass, and they and living nature, wouldn't be able to exist.
See also
References
- 1 2 3 Berger A, Crozier G, Bisogno T, Cavaliere P, Innis S, Di Marzo V (May 2001). "Anandamide and diet: inclusion of dietary arachidonate and docosahexaenoate leads to increased brain levels of the corresponding N-acylethanolamines in piglets". Proceedings of the National Academy of Sciences of the United States of America. 98 (11): 6402–6406. Bibcode:2001PNAS...98.6402B. doi:10.1073/pnas.101119098. PMC 33480. PMID 11353819.
- 1 2 Galasso I, Russo R, Mapelli S, Ponzoni E, Brambilla IM, Battelli G, Reggiani R (2016-05-20). "Variability in Seed Traits in a Collection of Cannabis sativa L. Genotypes". Frontiers in Plant Science. 7: 688. doi:10.3389/fpls.2016.00688. PMC 4873519. PMID 27242881.
- 1 2 Charytoniuk T, Zywno H, Berk K, Bzdega W, Kolakowski A, Chabowski A, Konstantynowicz-Nowicka K (March 2022). "The Endocannabinoid System and Physical Activity-A Robust Duo in the Novel Therapeutic Approach against Metabolic Disorders". International Journal of Molecular Sciences. 23 (6): 3083. doi:10.3390/ijms23063083. PMC 8948925. PMID 35328503.
- 1 2 3 Stone NL, Millar SA, Herrod PJ, Barrett DA, Ortori CA, Mellon VA, O'Sullivan SE (2018-11-26). "An Analysis of Endocannabinoid Concentrations and Mood Following Singing and Exercise in Healthy Volunteers". Frontiers in Behavioral Neuroscience. 12: 269. doi:10.3389/fnbeh.2018.00269. PMC 6275239. PMID 30534062.
 Material was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
Material was copied from this source, which is available under a Creative Commons Attribution 4.0 International License. - ↑ Blancaflor EB, Chapman KD (2006). "Similarities Between Endocannabinoid Signaling in Animal Systems and N-Acylethanolamine Metabolism in Plants". In Baluška F, Mancuso S, Volkmann D (eds.). Communication in Plants: Neuronal Aspects of Plant Life. Berlin, Heidelberg: Springer. pp. 205–219. doi:10.1007/978-3-540-28516-8_14. ISBN 978-3-540-28516-8.
- ↑ Fasano, Silvia; Meccariello, Rosaria; Cobellis, Gilda; Chianese, Rosanna; Cacciola, Giovanna; Chioccarelli, Teresa; Pierantoni, Riccardo (April 2009). "The Endocannabinoid System: An Ancient Signaling Involved in the Control of Male Fertility". Annals of the New York Academy of Sciences. 1163 (1): 112–124. Bibcode:2009NYASA1163..112F. doi:10.1111/j.1749-6632.2009.04437.x. PMID 19456333. S2CID 6304998.
- ↑ Elphick, M. R.; Egertová, M. (2005), Pertwee, Roger G. (ed.), "The Phylogenetic Distribution and Evolutionary Origins of Endocannabinoid Signalling", Cannabinoids, Handbook of Experimental Pharmacology, Berlin, Heidelberg: Springer, no. 168, pp. 283–297, doi:10.1007/3-540-26573-2_9, ISBN 978-3-540-26573-3, PMID 16596778
- ↑ McPartland, John M (2004-04-01). "Phylogenomic and chemotaxonomic analysis of the endocannabinoid system". Brain Research Reviews. 45 (1): 18–29. doi:10.1016/j.brainresrev.2003.11.005. ISSN 0165-0173. PMID 15063097. S2CID 25038370.
- ↑ Okamoto Y, Morishita J, Tsuboi K, Tonai T, Ueda N (February 2004). "Molecular characterization of a phospholipase D generating anandamide and its congeners". The Journal of Biological Chemistry. 279 (7): 5298–5305. doi:10.1074/jbc.M306642200. PMID 14634025.
- ↑ For example, note synonyms in PubChem for oleoylethanolamine.
- 1 2 Silva, Hugo-Henrique; Tavares, Valéria; Silva, Maria-Raquel G.; Neto, Beatriz Vieira; Cerqueira, Fátima; Medeiros, Rui (2023-03-26). "Association of FAAH rs324420 (C385A) Polymorphism with High-Level Performance in Volleyball Players". Genes. 14 (6): 1164. doi:10.3390/genes14061164. ISSN 2073-4425. PMC 10298391. PMID 37372343.
 This article incorporates text from this source, which is available under the CC BY 4.0 license.
This article incorporates text from this source, which is available under the CC BY 4.0 license. - ↑ Hayes, Alexander C. (2013). "Identification of N-acylethanolamines in Dictyostelium discoideum and confirmation of their hydrolysis by fatty acid amide hydrolase". J. Lipid Res. 54 (2): 457–466. doi:10.1194/jlr.M032219. PMC 3588872. PMID 23187822.
- 1 2 3 Shin, Kyong-Oh (2021-08-02). "N-Palmitoyl Serinol Stimulates Ceramide Production through a CB1-Dependent Mechanism in In Vitro Model of Skin Inflammation". Int J Mol Sci. 22 (15): 8302. doi:10.3390/ijms22158302. PMC 8348051. PMID 34361066. This article incorporates text from this source, which is available under the CC BY 4.0 license.
- ↑ The list and references provided are based on background discussion in Okamoto Y, Morishita J, Tsuboi K, Tonai T, Ueda N (February 2004). "Molecular characterization of a phospholipase D generating anandamide and its congeners". The Journal of Biological Chemistry. 279 (7): 5298–305. doi:10.1074/jbc.M306642200. PMID 14634025.
- ↑ PubChem. "Anandamide". pubchem.ncbi.nlm.nih.gov. Retrieved 2023-06-29.
- 1 2 Devane WA, Hanus L, Breuer A, Pertwee RG, Stevenson LA, Griffin G, et al. (December 1992). "Isolation and structure of a brain constituent that binds to the cannabinoid receptor". Science. 258 (5090): 1946–9. Bibcode:1992Sci...258.1946D. doi:10.1126/science.1470919. PMID 1470919.
- ↑ Di Marzo V (June 1998). "'Endocannabinoids' and other fatty acid derivatives with cannabimimetic properties: biochemistry and possible physiopathological relevance". Biochimica et Biophysica Acta (BBA) - Lipids and Lipid Metabolism. 1392 (2–3): 153–75. doi:10.1016/s0005-2760(98)00042-3. PMID 9630590.
- ↑ Di Marzo V, De Petrocellis L, Fezza F, Ligresti A, Bisogno T (2002). "Anandamide receptors". Prostaglandins, Leukotrienes, and Essential Fatty Acids. 66 (2–3): 377–91. doi:10.1054/plef.2001.0349. PMID 12052051.
- 1 2 Calignano A, La Rana G, Giuffrida A, Piomelli D (July 1998). "Control of pain initiation by endogenous cannabinoids". Nature. 394 (6690): 277–81. Bibcode:1998Natur.394..277C. doi:10.1038/28393. PMID 9685157. S2CID 4418082.
- 1 2 Maccarrone M, van der Stelt M, Rossi A, Veldink GA, Vliegenthart JF, Agrò AF (November 1998). "Anandamide hydrolysis by human cells in culture and brain". The Journal of Biological Chemistry. 273 (48): 32332–32339. doi:10.1074/jbc.273.48.32332. hdl:1887/50863. PMID 9822713.
- 1 2 Walter L, Franklin A, Witting A, Moller T, Stella N (June 2002). "Astrocytes in culture produce anandamide and other acylethanolamides". The Journal of Biological Chemistry. 277 (23): 20869–76. doi:10.1074/jbc.M110813200. PMID 11916961.
- 1 2 3 Kamlekar RK, Swamy MJ (July 2006). "Molecular packing and intermolecular interactions in two structural polymorphs of N-palmitoylethanolamine, a type 2 cannabinoid receptor agonist". Journal of Lipid Research. 47 (7): 1424–33. doi:10.1194/jlr.M600043-JLR200. PMID 16609146.
- ↑ Verme, Jesse Lo; Fu, Jin; Astarita, Giuseppe; Rana, Giovanna La; Russo, Roberto; Calignano, Antonio; Piomelli, Daniele (2005-01-01). "The Nuclear Receptor Peroxisome Proliferator-Activated Receptor-α Mediates the Anti-Inflammatory Actions of Palmitoylethanolamide". Molecular Pharmacology. 67 (1): 15–19. doi:10.1124/mol.104.006353. ISSN 0026-895X. PMID 15465922. S2CID 12671741.
- ↑ Lambert DM, Vandevoorde S, Jonsson KO, Fowler CJ (March 2002). "The palmitoylethanolamide family: a new class of anti-inflammatory agents?". Current Medicinal Chemistry. 9 (6): 663–74. doi:10.2174/0929867023370707. PMID 11945130.
- ↑ Rahman IA, Tsuboi K, Uyama T, Ueda N (August 2014). "New players in the fatty acyl ethanolamide metabolism". Pharmacological Research. Lipid amide signaling: regulation, physiological roles and pathological implications. 86: 1–10. doi:10.1016/j.phrs.2014.04.001. PMID 24747663.
- ↑ Darmani, Nissar A.; Izzo, Angelo A.; Degenhardt, Brian; Valenti, Marta; Scaglione, Giuseppe; Capasso, Raffaele; Sorrentini, Italo; Di Marzo, Vincenzo (2005-06-01). "Involvement of the cannabimimetic compound, N-palmitoyl-ethanolamine, in inflammatory and neuropathic conditions: Review of the available pre-clinical data, and first human studies". Neuropharmacology. Future Directions in Cannabinoid Therapeutics: “From the Bench to the Clinic”. 48 (8): 1154–1163. doi:10.1016/j.neuropharm.2005.01.001. ISSN 0028-3908. PMID 15910891. S2CID 14828175.
- 1 2 Chapman KD, Venables B, Markovic R, Bettinger C (August 1999). "N-Acylethanolamines in seeds. Quantification Of molecular species and their degradation upon imbibition". Plant Physiology. 120 (4): 1157–64. doi:10.1104/pp.120.4.1157. PMC 59349. PMID 10444099.
- ↑ Motes CM, Pechter P, Yoo CM, Wang YS, Chapman KD, Blancaflor EB (December 2005). "Differential effects of two phospholipase D inhibitors, 1-butanol and N-acylethanolamine, on in vivo cytoskeletal organization and Arabidopsis seedling growth". Protoplasma. 226 (3–4): 109–23. doi:10.1007/s00709-005-0124-4. PMID 16333570. S2CID 19838345.
- 1 2 PubChem. "Alpha-Linolenoyl ethanolamide". pubchem.ncbi.nlm.nih.gov. Retrieved 2023-08-03.
- 1 2 Jones, Peter J. H.; Lin, Lin; Gillingham, Leah G.; Yang, Haifeng; Omar, Jaclyn M. (2014-12-01). "Modulation of plasma N-acylethanolamine levels and physiological parameters by dietary fatty acid composition in humans". Journal of Lipid Research. 55 (12): 2655–2664. doi:10.1194/jlr.P051235. ISSN 0022-2275. PMC 4242457. PMID 25262934.
- 1 2 Rodríguez de Fonseca F, Navarro M, Gómez R, Escuredo L, Nava F, Fu J, et al. (November 2001). "An anorexic lipid mediator regulated by feeding". Nature. 414 (6860): 209–212. Bibcode:2001Natur.414..209R. doi:10.1038/35102582. PMID 11700558. S2CID 4430005.
- ↑ Kilaru A, Tamura P, Isaac G, Welti R, Venables BJ, Seier E, Chapman KD (September 2012). "Lipidomic analysis of N-acylphosphatidylethanolamine molecular species in Arabidopsis suggests feedback regulation by N-acylethanolamines". Planta. 236 (3): 809–824. Bibcode:2012Plant.236..809K. doi:10.1007/s00425-012-1669-z. PMC 3579225. PMID 22673881.
- ↑ Di Marzo V, Piscitelli F (May 2011). "Gut feelings about the endocannabinoid system". Neurogastroenterology and Motility. 23 (5): 391–398. doi:10.1111/j.1365-2982.2011.01689.x. PMID 21481098.
- 1 2 3 4 5 Tripathy S, Kleppinger-Sparace K, Dixon RA, Chapman KD (April 2003). "N-acylethanolamine signaling in tobacco is mediated by a membrane-associated, high-affinity binding protein". Plant Physiology. 131 (4): 1781–91. doi:10.1104/pp.102.014936. PMC 166934. PMID 12692337.
- ↑ Hofmann U, Domeier E, Frantz S, Laser M, Weckler B, Kuhlencordt P, et al. (June 2003). "Increased myocardial oxygen consumption by TNF-alpha is mediated by a sphingosine signaling pathway". American Journal of Physiology. Heart and Circulatory Physiology. 284 (6): H2100-5. doi:10.1152/ajpheart.00888.2002. PMID 12560208.
- ↑ Amadou A, Nawrocki A, Best-Belpomme M, Pavoine C, Pecker F (June 2002). "Arachidonic acid mediates dual effect of TNF-alpha on Ca2+ transients and contraction of adult rat cardiomyocytes". American Journal of Physiology. Cell Physiology. 282 (6): C1339-47. doi:10.1152/ajpcell.00471.2001. PMID 11997249.
- ↑ "Linoleoyl ethanolamide". PubChem. U.S. National Library of Medicine. Retrieved 2022-10-29.
- 1 2 "Docosahexaenoyl Ethanolamide". PubChem. U.S. National Library of Medicine. Retrieved 2022-11-19.
- 1 2 3 Balvers MG, Verhoeckx KC, Plastina P, Wortelboer HM, Meijerink J, Witkamp RF (October 2010). "Docosahexaenoic acid and eicosapentaenoic acid are converted by 3T3-L1 adipocytes to N-acyl ethanolamines with anti-inflammatory properties". Biochimica et Biophysica Acta (BBA) - Molecular and Cell Biology of Lipids. 1801 (10): 1107–14. doi:10.1016/j.bbalip.2010.06.006. PMID 20601112.
- 1 2 3 Brown, Iain; Cascio, Maria G.; Rotondo, Dino; Pertwee, Roger G.; Heys, Steven D.; Wahle, Klaus W. J. (2013-01-01). "Cannabinoids and omega-3/6 endocannabinoids as cell death and anticancer modulators". Progress in Lipid Research. 52 (1): 80–109. doi:10.1016/j.plipres.2012.10.001. ISSN 0163-7827. PMID 23103355.
- ↑ Brown I, Cascio MG, Wahle KW, Smoum R, Mechoulam R, Ross RA, et al. (September 2010). "Cannabinoid receptor-dependent and -independent anti-proliferative effects of omega-3 ethanolamides in androgen receptor-positive and -negative prostate cancer cell lines". Carcinogenesis. 31 (9): 1584–91. doi:10.1093/carcin/bgq151. PMC 2930808. PMID 20660502.
- ↑ Kim HY, Spector AA, Xiong ZM (November 2011). "A synaptogenic amide N-docosahexaenoylethanolamide promotes hippocampal development". Prostaglandins & Other Lipid Mediators. 96 (1–4): 114–20. doi:10.1016/j.prostaglandins.2011.07.002. PMC 3215906. PMID 21810478.
- ↑ Lee JW, Huang BX, Kwon H, Rashid MA, Kharebava G, Desai A, et al. (October 2016). "Orphan GPR110 (ADGRF1) targeted by N-docosahexaenoylethanolamine in development of neurons and cognitive function". Nature Communications. 7 (1): 13123. Bibcode:2016NatCo...713123L. doi:10.1038/ncomms13123. PMC 5075789. PMID 27759003.
- 1 2 PubChem. "Eicosapentaenoyl Ethanolamide". pubchem.ncbi.nlm.nih.gov. Retrieved 2023-08-05.
- ↑ Walter L, Franklin A, Witting A, Wade C, Xie Y, Kunos G, et al. (February 2003). "Nonpsychotropic cannabinoid receptors regulate microglial cell migration". The Journal of Neuroscience. 23 (4): 1398–405. doi:10.1523/JNEUROSCI.23-04-01398.2003. PMC 6742252. PMID 12598628.
- ↑ Magotti P, Bauer I, Igarashi M, Babagoli M, Marotta R, Piomelli D, Garau G (March 2015). "Structure of human N-acylphosphatidylethanolamine-hydrolyzing phospholipase D: regulation of fatty acid ethanolamide biosynthesis by bile acids". Structure. 23 (3): 598–604. doi:10.1016/j.str.2014.12.018. PMC 4351732. PMID 25684574.
- 1 2 Mock ED, Mustafa M, Gunduz-Cinar O, Cinar R, Petrie GN, Kantae V, et al. (June 2020). "Discovery of a NAPE-PLD inhibitor that modulates emotional behavior in mice". Nature Chemical Biology. 16 (6): 667–675. doi:10.1038/s41589-020-0528-7. PMC 7468568. PMID 32393901.
- ↑ Tsuboi K, Okamoto Y, Ikematsu N, Inoue M, Shimizu Y, Uyama T, et al. (October 2011). "Enzymatic formation of N-acylethanolamines from N-acylethanolamine plasmalogen through N-acylphosphatidylethanolamine-hydrolyzing phospholipase D-dependent and -independent pathways". Biochimica et Biophysica Acta (BBA) - Molecular and Cell Biology of Lipids. 1811 (10): 565–577. doi:10.1016/j.bbalip.2011.07.009. PMID 21801852.
- ↑ Wellner N, Diep TA, Janfelt C, Hansen HS (March 2013). "N-acylation of phosphatidylethanolamine and its biological functions in mammals". Biochimica et Biophysica Acta (BBA) - Molecular and Cell Biology of Lipids. 1831 (3): 652–662. doi:10.1016/j.bbalip.2012.08.019. PMID 23000428.
- ↑ Serrano A, Parsons LH (December 2011). "Endocannabinoid influence in drug reinforcement, dependence and addiction-related behaviors". Pharmacology & Therapeutics. 132 (3): 215–241. doi:10.1016/j.pharmthera.2011.06.005. PMC 3209522. PMID 21798285.
- 1 2 Oka, Saori (2006). "Involvement of the Cannabinoid CB2 Receptor and Its Endogenous Ligand 2-Arachidonoylglycerol in Oxazolone-Induced Contact Dermatitis in Mice". Journal of Immunology. 177 (12): 8796–9505. doi:10.4049/jimmunol.177.12.8796. PMID 17142782. S2CID 11946479. Retrieved 2023-03-24.
- 1 2 McPartland JM, Guy GW, Di Marzo V (2014-03-12). "Care and feeding of the endocannabinoid system: a systematic review of potential clinical interventions that upregulate the endocannabinoid system". PLOS ONE. 9 (3): e89566. Bibcode:2014PLoSO...989566M. doi:10.1371/journal.pone.0089566. PMC 3951193. PMID 24622769. Text was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
- ↑ Di Marzo V, Silvestri C (August 2019). "Lifestyle and Metabolic Syndrome: Contribution of the Endocannabinoidome". Nutrients. 11 (8). 1956. doi:10.3390/nu11081956. PMC 6722643. PMID 31434293.
- 1 2 Surowiec I, Gouveia-Figueira S, Orikiiriza J, Lindquist E, Bonde M, Magambo J, et al. (September 2017). "The oxylipin and endocannabidome responses in acute phase Plasmodium falciparum malaria in children". Malaria Journal. 16 (1): 358. doi:10.1186/s12936-017-2001-y. PMC 5591560. PMID 28886714.
- ↑ Lacroix S (2020). "Dietary fatty acid intake and gut microbiota determine circulating endocannabinoidome signaling beyond the efect of body fat" (PDF). Nature research.
- ↑ Manca, Claudia (2020). "Germ-free mice exhibit profound gut microbiota-dependent alterations of intestinal endocannabinoidome signaling". J Lipid Res. 61 (1): 70–85. doi:10.1194/jlr.RA119000424. PMC 6939599. PMID 31690638.
- ↑ Venneri, Tommaso (2023-05-15). "Altered endocannabinoidome bioactive lipid levels accompany reduced DNBS-induced colonic inflammation in germ-free mice". Lipids in Health and Disease. 22 (1): 63. doi:10.1186/s12944-023-01823-1. PMC 10186685. PMID 37189092.
- ↑ The Scientist, archived from the original on 2021-12-15, retrieved 2021-03-14
- ↑ Blancaflor EB, Kilaru A, Keereetaweep J, Khan BR, Faure L, Chapman KD (August 2014). "N-Acylethanolamines: lipid metabolites with functions in plant growth and development". The Plant Journal. 79 (4): 568–83. doi:10.1111/tpj.12427. PMID 24397856.
- 1 2 3 Sorrentino G (March 2021). "Introduction to emerging industrial applications of cannabis (Cannabis sativa L.)". Rendiconti Lincei. Scienze Fisiche e Naturali. 32 (2): 233–243. doi:10.1007/s12210-021-00979-1. PMC 7978457. PMID 33777341.
- 1 2 Rockström J, Steffen W, Noone K, Persson Å, Chapin III FS, Lambin E, et al. (2009-11-18). "Planetary Boundaries: Exploring the Safe Operating Space for Humanity". Ecology and Society. 14 (2). doi:10.5751/ES-03180-140232. hdl:10535/5421. ISSN 1708-3087.
- ↑ Soderstrom K (2009). "Lessons from Nonmammalian Species". In Kendall D, Alexander S (eds.). Behavioral Neurobiology of the Endocannabinoid System. Current Topics in Behavioral Neurosciences. Vol. 1. Berlin, Heidelberg: Springer. pp. 173–98. doi:10.1007/978-3-540-88955-7_7. ISBN 978-3-540-88955-7. PMID 21104384.
- ↑ DeVries MS, Cordes MA, Rodriguez JD, Stevenson SA, Riters LV (August 2016). "Neural endocannabinoid CB1 receptor expression, social status, and behavior in male European starlings". Brain Research. 1644: 240–8. doi:10.1016/j.brainres.2016.05.031. PMC 4904780. PMID 27206544.
- 1 2 Melamede R (September 2005). "Harm reduction--the cannabis paradox". Harm Reduction Journal. 2 (1): 17. doi:10.1186/1477-7517-2-17. PMC 1261530. PMID 16179090. Text was copied from this source, which is available under a Creative Commons Attribution 2.0 Generic (CC BY 2.0) license.
- ↑ Wang H, Matsumoto H, Guo Y, Paria BC, Roberts RL, Dey SK (December 2003). "Differential G protein-coupled cannabinoid receptor signaling by anandamide directs blastocyst activation for implantation". Proceedings of the National Academy of Sciences of the United States of America. 100 (25): 14914–14919. Bibcode:2003PNAS..10014914W. doi:10.1073/pnas.2436379100. PMC 299852. PMID 14645706.
- ↑ Wang H, Xie H, Dey SK (2006). "Endocannabinoid signaling directs periimplantation events". The AAPS Journal. 8 (2): E425–E432. doi:10.1007/BF02854916. PMC 3231559. PMID 16808046.
- 1 2 3 Fride E (October 2004). "The endocannabinoid-CB(1) receptor system in pre- and postnatal life". European Journal of Pharmacology. SPECIAL CELEBRATORY VOLUME 500 Dedicated to Professor David de Wied Honorary and Founding Editor. 500 (1–3): 289–297. doi:10.1016/j.ejphar.2004.07.033. PMID 15464041.
- 1 2 Trabucco E, Acone G, Marenna A, Pierantoni R, Cacciola G, Chioccarelli T, et al. (June 2009). "Endocannabinoid system in first trimester placenta: low FAAH and high CB1 expression characterize spontaneous miscarriage". Placenta. 30 (6): 516–522. doi:10.1016/j.placenta.2009.03.015. PMID 19419760.
- ↑ Maccarrone M, Valensise H, Bari M, Lazzarin N, Romanini C, Finazzi-Agrò A (April 2000). "Relation between decreased anandamide hydrolase concentrations in human lymphocytes and miscarriage". Lancet. 355 (9212): 1326–1329. doi:10.1016/S0140-6736(00)02115-2. PMID 10776746. S2CID 39733100.
- ↑ Wang H, Guo Y, Wang D, Kingsley PJ, Marnett LJ, Das SK, et al. (October 2004). "Aberrant cannabinoid signaling impairs oviductal transport of embryos". Nature Medicine. 10 (10): 1074–80. doi:10.1038/nm1104. PMID 15378054. S2CID 29106466.
- ↑ Stadel, Rebecca; Ahn, Kwang H.; Kendall, Debra A. (2011). "The Cannabinoid Type-1 Receptor Carboxyl-Terminus, More Than Just a Tail". Journal of Neurochemistry. 117 (1): 1–18. doi:10.1111/j.1471-4159.2011.07186.x. ISSN 0022-3042. PMC 3055937. PMID 21244428.
- ↑ Pertwee RG (January 2008). "The diverse CB1 and CB2 receptor pharmacology of three plant cannabinoids: delta9-tetrahydrocannabinol, cannabidiol and delta9-tetrahydrocannabivarin". British Journal of Pharmacology. 153 (2): 199–215. doi:10.1038/sj.bjp.0707442. PMC 2219532. PMID 17828291.
- ↑ Morales P, Hurst DP, Reggio PH (2017). "Molecular Targets of the Phytocannabinoids: A Complex Picture". Phytocannabinoids. Progress in the Chemistry of Organic Natural Products. Vol. 103. Cham: Springer. pp. 103–131. doi:10.1007/978-3-319-45541-9_4. ISBN 978-3-319-45539-6. PMC 5345356. PMID 28120232.
- 1 2 Thieme U, Schelling G, Hauer D, Greif R, Dame T, Laubender RP, et al. (2014). "Quantification of anandamide and 2-arachidonoylglycerol plasma levels to examine potential influences of tetrahydrocannabinol application on the endocannabinoid system in humans". Drug Testing and Analysis. 6 (1–2): 17–23. doi:10.1002/dta.1561. PMID 24424856.
- 1 2 Xiong, Wei (2011). "Cannabinoid potentiation of glycine receptors contributes to cannabis-induced analgesia". Nature Chemical Biology. 7 (5): 296–303. doi:10.1038/nchembio.552. PMC 3388539. PMID 21460829.
- ↑ O'Sullivan, S E (2007). "Cannabinoids go nuclear: evidence for activation of peroxisome proliferator-activated receptors". British Journal of Pharmacology. 152 (5): 576–582. doi:10.1038/sj.bjp.0707423. ISSN 0007-1188. PMC 2190029. PMID 17704824.
- ↑ Roloff AM, Thayer SA (April 2009). "Modulation of excitatory synaptic transmission by Delta 9-tetrahydrocannabinol switches from agonist to antagonist depending on firing rate". Molecular Pharmacology. 75 (4): 892–900. doi:10.1124/mol.108.051482. PMC 2684930. PMID 19118122.
- ↑ Lipina, Christopher (2017-01-24). "The endocannabinoid system: 'NO' longer anonymous in the control of nitrergic signalling?". J Mol Cell Biol. 9 (2): 91–103. doi:10.1093/jmcb/mjx008. PMC 5439392. PMID 28130308.
- 1 2 Shmist, Yelena A.; Goncharov, Igor; Eichler, Maor; Shneyvays, Vladimir; Isaac, Ahuva; Vogel, Zvi; Shainberg, Asher (2006-02-01). "Delta-9-tetrahydrocannabinol protects cardiac cells from hypoxia via CB2 receptor activation and nitric oxide production". Molecular and Cellular Biochemistry. 283 (1): 75–83. doi:10.1007/s11010-006-2346-y. ISSN 1573-4919. PMID 16444588. S2CID 24074568.
- ↑ Dennedy MC, Friel AM, Houlihan DD, Broderick VM, Smith T, Morrison JJ (January 2004). "Cannabinoids and the human uterus during pregnancy" (PDF). American Journal of Obstetrics and Gynecology. 190 (1): 2–9, discussion 3A. doi:10.1016/j.ajog.2003.07.013. PMID 14749627.
- ↑ Datta P, Melkus MW, Rewers-Felkins K, Patel D, Bateman T, Baker T, Hale TW (July 2021). "Human Milk Endocannabinoid Levels as a Function of Obesity and Diurnal Rhythm". Nutrients. 13 (7): 2297. doi:10.3390/nu13072297. PMC 8308542. PMID 34371807.
- ↑ Fride E, Suris R, Weidenfeld J, Mechoulam R (September 2005). "Differential response to acute and repeated stress in cannabinoid CB1 receptor knockout newborn and adult mice". Behavioural Pharmacology. 16 (5–6): 431–440. doi:10.1097/00008877-200509000-00016. PMID 16148448. S2CID 25548954.
- ↑ Botsford, Chloe; Brellenthin, Angelique G.; Cisler, Josh M.; Hillard, Cecilia J.; Koltyn, Kelli F.; Crombie, Kevin M. (2023-01-01). "Circulating endocannabinoids and psychological outcomes in women with PTSD". Journal of Anxiety Disorders. 93: 102656. doi:10.1016/j.janxdis.2022.102656. ISSN 0887-6185. PMC 9839585. PMID 36469982.
- 1 2 Hill, Matthew N. (2009). "Circulating Endocannabinoids and N-Acyl Ethanolamines Are Differentially Regulated in Major Depression and Following Exposure to Social Stress". Psychoneuroendocrinology. 34 (8): 1257–1262. doi:10.1016/j.psyneuen.2009.03.013. PMC 2716432. PMID 19394765.
- ↑ Wingenfeld, Katja; Dettenborn, Lucia; Kirschbaum, Clemens; Gao, Wei; Otte, Christian; Roepke, Stefan (2018-07-04). "Reduced levels of the endocannabinoid arachidonylethanolamide (AEA) in hair in patients with borderline personality disorder – a pilot study". Stress. 21 (4): 366–369. doi:10.1080/10253890.2018.1451837. ISSN 1025-3890. PMID 29546791. S2CID 3929753.
- 1 2 Crippa, José; Zuardi, Antonio; Hallak, Jaime (2010). "Therapeutical use of the cannabinoids in psychiatry". Braz J Psychiatry. 32: 56–66. PMID 20512271 – via PubMed Central.
- ↑ Cousens, Kenneth; DiMascio, Alberto (1973-12-01). "(−)δ9 THC as an hypnotic". Psychopharmacologia. 33 (4): 355–364. doi:10.1007/BF00437513. ISSN 1432-2072. PMID 4776660. S2CID 8775767.
- 1 2 Kolla, Nathan J. (2020-06-10). "Elevated fatty acid amide hydrolase in the prefrontal cortex of borderline personality disorder: a [11C]CURB positron emission tomography study". Neuropsychopharmacology. 45 (11): 1834–1841. doi:10.1038/s41386-020-0731-y. PMC 7608329. PMID 32521537.
- 1 2 Wei D, Lee D, Cox CD, Karsten CA, Peñagarikano O, Geschwind DH, et al. (November 2015). "Endocannabinoid signaling mediates oxytocin-driven social reward". Proceedings of the National Academy of Sciences of the United States of America. 112 (45): 14084–14089. Bibcode:2015PNAS..11214084W. doi:10.1073/pnas.1509795112. PMC 4653148. PMID 26504214.
- ↑ Ritter, Catherine; Broers, Barbara; Elger, Bernice S. (2013-11-01). "Cannabis use in a Swiss male prison: Qualitative study exploring detainees' and staffs' perspectives". International Journal of Drug Policy. 24 (6): 573–578. doi:10.1016/j.drugpo.2013.05.001. ISSN 0955-3959. PMID 23773686.
- ↑ Jiménez-Vinaja, R. (2022-03-31). "Jailhouse rock: Health status, patterns of use, and subjective benefits of cannabis use in a sample of inmates in Spanish prisons". Rev Esp Sanid Penit. 24 (1): 9–14. doi:10.18176/resp.00044. PMC 9017606. PMID 35411912.
- ↑ Excerpt: The House I Live In - The Chain of Destruction, retrieved 2023-08-15
- ↑ Kolind, Torsten; Frank, Vibeke Asmussen; Dahl, Helle (2010-01-01). "Drug treatment or alleviating the negative consequences of imprisonment? A critical view of prison-based drug treatment in Denmark". International Journal of Drug Policy. 21 (1): 43–48. doi:10.1016/j.drugpo.2009.03.002. ISSN 0955-3959. PMID 19427186.
- ↑ de Lima e Silva Surjus, Luciana Togni (2023-04-05). "Use of Marijuana to Promote Well-Being: Effects of Use and Prohibition in the Daily Lives of Brazilian Adults". Subst Abuse. 17. doi:10.1177/11782218231162469. PMC 10084564. PMID 37051015.
- 1 2 El-Alfy, Abir T. (2010-03-21). "Antidepressant-like effect of Δ9-tetrahydrocannabinol and other cannabinoids isolated from Cannabis sativa L". Pharmacol Biochem Behav. 95 (4): 434–442. doi:10.1016/j.pbb.2010.03.004. PMC 2866040. PMID 20332000.
- ↑ His Majesty's, Government of Nepal (1975-08-08). "CHAPTER VI NARCOTIC DRUGS AND PSYCHOTROPIC SUBSTANCES - Declarations and Reservations for Nepal". un.org.
- ↑ Hamayun, Muhammad; Shinwari, Zabta Khan (2004-12-22). "Folk Methodology of Charas (Hashish) Production and Its Marketing at Afridi Tirah, Federally Administered Tribal Areas (FATA), Pakistan". Journal of Industrial Hemp. 9 (2): 41–50. doi:10.1300/J237v09n02_04. ISSN 1537-7881. S2CID 85423253.
- ↑ "The Science of Marijuana, 2nd edn". British Journal of Clinical Pharmacology. 67 (2): 268. 2009-02-23. doi:10.1111/j.1365-2125.2008.03355.x. PMC 2670388.
- ↑ Adinoff, Bryon; Reiman, Amanda (2019-11-02). "Implementing social justice in the transition from illicit to legal cannabis". The American Journal of Drug and Alcohol Abuse. 45 (6): 673–688. doi:10.1080/00952990.2019.1674862. ISSN 0095-2990. PMID 31634005. S2CID 204834660.
- ↑ Iōakeimoglou, Geōrgios; Wolstenholme, G. E. W.; Knight, Julie (1965). "Hashish its chemistry and pharmacology. In honor of Professor Dr. G. Joachimoglu | WorldCat.org". www.worldcat.org. Retrieved 2023-08-13.
- ↑ Morris, Robert G.; TenEyck, Michael; Barnes, J. C.; Kovandzic, Tomislav V. (2014-03-26). "The Effect of Medical Marijuana Laws on Crime: Evidence from State Panel Data, 1990-2006". PLOS ONE. 9 (3): e92816. Bibcode:2014PLoSO...992816M. doi:10.1371/journal.pone.0092816. PMC 3966811. PMID 24671103.
- 1 2 Anderson DM, Rees DI, Sabia JJ (January 2012). "High on Life? Medical Marijuana Laws and Suicide" (PDF). Institute for the Study of Labor (IZA).
- ↑ Russo, Ethan B. (2023-01-02). "When The Pharmacopoeia Fails: Cannabis for Pain". Pain Medicine (Malden, Mass.). 24 (7): 912–913. doi:10.1093/pm/pnac208. PMID 36592031. Retrieved 2023-08-13.
- ↑ Bojesen, Bo (1979-04-06). "Title: Tremmeværk/latticework - Situation: Prisoner in solitary confinement gets a visit, where the official visits the prisoner, who sits alone in the dark - Subtitle: The Human Rights Commission must now take a closer look at the long-term isolation of Danish remand prisoners.- Dialogue: You can try to talk to him, but he's been so strangely withdrawn over the last seven months. - Digitale samlinger". www5.kb.dk. Retrieved 2023-08-13.
- ↑ Subramanian C, Frank MW, Batte JL, Whaley SG, Rock CO (June 2019). "Oleate hydratase from Staphylococcus aureus protects against palmitoleic acid, the major antimicrobial fatty acid produced by mammalian skin". The Journal of Biological Chemistry. 294 (23): 9285–9294. doi:10.1074/jbc.RA119.008439. PMC 6556589. PMID 31018965.
- ↑ Vanhoutte K, Vanhoutte J (2009-06-22). "Composition comprising omega-7 and/or omega-4 fatty acids". Google Patents.
- ↑ Compton DL, Laszlo JA, Appell M, Vermillion KE, Evans KO (2012). "Influence of Fatty Acid Desaturation on Spontaneous Acyl Migration in 2-Monoacylglycerols". Journal of the American Oil Chemists' Society. 89 (12): 2259–2267. doi:10.1007/s11746-012-2113-z. ISSN 1558-9331. S2CID 83629022.
- ↑ Manca C, Carta G, Murru E, Abolghasemi A, Ansar H, Errigo A, et al. (March 2021). "Circulating fatty acids and endocannabinoidome-related mediator profiles associated to human longevity". GeroScience. 43 (4): 1783–1798. doi:10.1007/s11357-021-00342-0. PMC 8492808. PMID 33650014.
- ↑ Simopoulos AP (October 2002). "The importance of the ratio of omega-6/omega-3 essential fatty acids". Biomedicine & Pharmacotherapy. 56 (8): 365–379. doi:10.1016/S0753-3322(02)00253-6. PMID 12442909.
- ↑ Ryz, Natasha R.; Remillard, David J.; Russo, Ethan B. (2017-08-01). "Cannabis Roots: A Traditional Therapy with Future Potential for Treating Inflammation and Pain". Cannabis and Cannabinoid Research. 2 (1): 210–216. doi:10.1089/can.2017.0028. ISSN 2378-8763. PMC 5628559. PMID 29082318.
- ↑ PubChem. "Ergost-5-en-3-ol". pubchem.ncbi.nlm.nih.gov. Retrieved 2023-03-26.
- ↑ PubChem. "p-Coumaroyltyramine". pubchem.ncbi.nlm.nih.gov. Retrieved 2023-03-26.
- ↑ Elhendawy, Mostafa A.; Wanas, Amira S.; Radwan, Mohamed M.; Azzaz, Nabil A.; Toson, ElShahat S.; ElSohly, Mahmoud A. (2018). "Chemical and Biological Studies of Cannabis sativa Roots". Medical Cannabis and Cannabinoids. 1 (2): 104–111. doi:10.1159/000495582. ISSN 2504-3889. PMC 7158750. PMID 32296742.
- ↑ Iftikhar, Amna (2021-12-20). "Applications of Cannabis Sativa L. in Food and Its Therapeutic Potential: From a Prohibited Drug to a Nutritional Supplement". Molecules. 26 (24): 7699. doi:10.3390/molecules26247699. PMC 8708517. PMID 34946779.
- ↑ "Hemp Seed Protein". Innvista. Retrieved 2022-10-31.
- ↑ "Omega Oils". Innvista. Retrieved 2022-10-31.
- ↑ "Hampefrøolie". www.nyborggaard.dk (in Danish). Retrieved 2023-07-22.
- ↑ "Hampeprotein". www.nyborggaard.dk (in Danish). Retrieved 2023-08-20.
- ↑ Guil-Guerrero JL, Rincón-Cervera MÁ, Venegas-Venegas E (2010). "Gamma-linolenic and stearidonic acids: Purification and upgrading of C18-PUFA oils". European Journal of Lipid Science and Technology. 112 (10): 1068–1081. doi:10.1002/ejlt.200900294. ISSN 1438-7697.
- ↑ Tóth, Kinga Fanni; Ádám, Dorottya; Bíró, Tamás; Oláh, Attila (2019-03-06). "Cannabinoid Signaling in the Skin: Therapeutic Potential of the "C(ut)annabinoid" System". Molecules. 24 (5): 918. doi:10.3390/molecules24050918. ISSN 1420-3049. PMC 6429381. PMID 30845666.
- ↑ Kendall, Alexandra C.; Pilkington, Suzanne M.; Massey, Karen A.; Sassano, Gary; Rhodes, Lesley E.; Nicolaou, Anna (2015-06-01). "Distribution of Bioactive Lipid Mediators in Human Skin". Journal of Investigative Dermatology. 135 (6): 1510–1520. doi:10.1038/jid.2015.41. ISSN 0022-202X. PMID 25668241. S2CID 2385238.
- 1 2 3 4 Courtney W (2008). "Marijuana Symposium Mendocino Sept 11 '08". YouTube. Archived from the original on 2021-12-15.
- ↑ PubChem. "(6aR,10aR)-1-Hydroxy-6,6,9-trimethyl-3-pentyl-6a,7,8,10a-tetrahydro-6H-dibenzo[b,d]pyran-4-carboxylic acid". pubchem.ncbi.nlm.nih.gov. Retrieved 2023-03-24.
- ↑ PubChem. "Cannabidiolic acid". pubchem.ncbi.nlm.nih.gov. Retrieved 2023-03-24.
- ↑ Nadal, Xavier; del Río, Carmen; Casano, Salvatore; Palomares, Belén; Ferreiro-Vera, Carlos; Navarrete, Carmen; Sánchez-Carnerero, Carolina; Cantarero, Irene; Bellido, Maria Luz; Meyer, Stefan; Morello, Gaetano; Appendino, Giovanni; Muñoz, Eduardo (December 2017). "Tetrahydrocannabinolic acid is a potent PPARγ agonist with neuroprotective activity". British Journal of Pharmacology. 174 (23): 4263–4276. doi:10.1111/bph.14019. ISSN 0007-1188. PMC 5731255. PMID 28853159.
- ↑ Petrosino, Stefania; Verde, Roberta; Vaia, Massimo; Allarà, Marco; Iuvone, Teresa; Marzo, Vincenzo Di (2018-06-01). "Anti-inflammatory Properties of Cannabidiol, a Nonpsychotropic Cannabinoid, in Experimental Allergic Contact Dermatitis". Journal of Pharmacology and Experimental Therapeutics. 365 (3): 652–663. doi:10.1124/jpet.117.244368. ISSN 0022-3565. PMID 29632236. S2CID 4739468.
- ↑ Karsak, Meliha (2007-06-08). "Attenuation of Allergic Contact Dermatitis Through the Endocannabinoid System". Science. 316 (5830): 1494–1497. Bibcode:2007Sci...316.1494K. doi:10.1126/science.1142265. PMID 17556587. S2CID 37611370.
- ↑ "8,11,14,17-Eicosatetraenoic acid". PubChem. U.S. National Library of Medicine. Retrieved 2022-11-28.
- ↑ Abedi E, Sahari MA (September 2014). "Long-chain polyunsaturated fatty acid sources and evaluation of their nutritional and functional properties". Food Science & Nutrition. 2 (5): 443–463. doi:10.1002/fsn3.121. PMC 4237475. PMID 25473503.
- ↑ "Anandamide". PubChem. U.S. National Library of Medicine. Retrieved 2022-11-28.
- 1 2 Sugiura T, Kondo S, Kishimoto S, Miyashita T, Nakane S, Kodaka T, et al. (January 2000). "Evidence that 2-arachidonoylglycerol but not N-palmitoylethanolamine or anandamide is the physiological ligand for the cannabinoid CB2 receptor. Comparison of the agonistic activities of various cannabinoid receptor ligands in HL-60 cells". The Journal of Biological Chemistry. 275 (1): 605–612. doi:10.1074/jbc.275.1.605. PMID 10617657.
- ↑ "2-Arachidonoylglycerol". PubChem. U.S. National Library of Medicine. Retrieved 2022-11-28.
- ↑ PubChem. "delta9-Tetrahydrocannabinolic acid". pubchem.ncbi.nlm.nih.gov. Retrieved 2023-08-05.
- 1 2 PubChem. "Dronabinol". pubchem.ncbi.nlm.nih.gov. Retrieved 2023-08-05.
- ↑ Guzmán, Manuel (2003). "Cannabinoids: potential anticancer agents". Nature Reviews Cancer. 3 (10): 745–755. doi:10.1038/nrc1188. ISSN 1474-1768. PMID 14570037. S2CID 7868655.
- ↑ Guzmán, Manuel; Sánchez, Cristina; Galve-Roperh, Ismael (2001-01-01). "Control of the cell survival/death decision by cannabinoids". Journal of Molecular Medicine. 78 (11): 613–625. doi:10.1007/s001090000177. ISSN 1432-1440. PMID 11269508. S2CID 12278192.
- ↑ Munson, A. E.; Harris, L. S.; Friedman, M. A.; Dewey, W. L.; Carchman, R. A. (1975-09-01). "Antineoplastic Activity of Cannabinoids". Journal of the National Cancer Institute. 55 (3): 597–602. doi:10.1093/jnci/55.3.597. PMID 1159836.
- 1 2 ElTelbany, Ahmed (2022-04-28). "Lower Rates of Hepatocellular Carcinoma Observed Among Cannabis Users: A Population-Based Study". Cureus. 14 (4): e24576. doi:10.7759/cureus.24576. PMC 9138632. PMID 35651376.
- ↑ Clark, Thomas M. (2021-10-13). "Scoping Review and Meta-Analysis Suggests that Cannabis Use May Reduce Cancer Risk in the United States". Cannabis Cannabinoid Res. 6 (5): 413–434. doi:10.1089/can.2019.0095. PMC 8612444. PMID 33998861.
- 1 2 Dyall SC (November 2017). "Interplay Between n-3 and n-6 Long-Chain Polyunsaturated Fatty Acids and the Endocannabinoid System in Brain Protection and Repair". Lipids. 52 (11): 885–900. doi:10.1007/s11745-017-4292-8. PMC 5656721. PMID 28875399.
- ↑ Gaitán AV, Wood JT, Solomons NW, Donohue JA, Ji L, Liu Y, et al. (June 2019). "Endocannabinoid Metabolome Characterization of Milk from Guatemalan Women Living in the Western Highlands". Current Developments in Nutrition. 3 (6): nzz018. doi:10.1093/cdn/nzz018. PMC 6517780. PMID 31111118.
- 1 2 Keereetaweep J, Chapman KD (2016). "Lipidomic Analysis of Endocannabinoid Signaling: Targeted Metabolite Identification and Quantification". Neural Plasticity. 2016: 2426398. doi:10.1155/2016/2426398. PMC 4709765. PMID 26839710.
- ↑ Yu M, Ives D, Ramesha CS (August 1997). "Synthesis of prostaglandin E2 ethanolamide from anandamide by cyclooxygenase-2". The Journal of Biological Chemistry. 272 (34): 21181–6. doi:10.1074/jbc.272.34.21181. PMID 9261124.
- ↑ Patsos HA, Hicks DJ, Dobson RR, Greenhough A, Woodman N, Lane JD, et al. (December 2005). "The endogenous cannabinoid, anandamide, induces cell death in colorectal carcinoma cells: a possible role for cyclooxygenase 2". Gut. 54 (12): 1741–50. doi:10.1136/gut.2005.073403. PMC 1774787. PMID 16099783.
- ↑ Ritter JK, Li C, Xia M, Poklis JL, Lichtman AH, Abdullah RA, et al. (September 2012). "Production and actions of the anandamide metabolite prostamide E2 in the renal medulla". The Journal of Pharmacology and Experimental Therapeutics. 342 (3): 770–9. doi:10.1124/jpet.112.196451. PMC 3422528. PMID 22685343.
- ↑ Touriño C, Oveisi F, Lockney J, Piomelli D, Maldonado R (March 2010). "FAAH deficiency promotes energy storage and enhances the motivation for food". International Journal of Obesity. 34 (3): 557–568. doi:10.1038/ijo.2009.262. PMC 3709605. PMID 20029375.
- ↑ Filipiuc LE, Ababei DC, Alexa-Stratulat T, Pricope CV, Bild V, Stefanescu R, et al. (November 2021). "Major Phytocannabinoids and Their Related Compounds: Should We Only Search for Drugs That Act on Cannabinoid Receptors?". Pharmaceutics. 13 (11): 1823. doi:10.3390/pharmaceutics13111823. PMC 8625816. PMID 34834237.
- 1 2 Weis F, Beiras-Fernandez A, Hauer D, Hornuss C, Sodian R, Kreth S, et al. (August 2010). "Effect of anaesthesia and cardiopulmonary bypass on blood endocannabinoid concentrations during cardiac surgery". British Journal of Anaesthesia. 105 (2): 139–144. doi:10.1093/bja/aeq117. PMID 20525978.
- 1 2 Murdolo G, Kempf K, Hammarstedt A, Herder C, Smith U, Jansson PA (September 2007). "Insulin differentially modulates the peripheral endocannabinoid system in human subcutaneous abdominal adipose tissue from lean and obese individuals". Journal of Endocrinological Investigation. 30 (8): RC17–RC21. doi:10.1007/BF03347440. PMID 17923791. S2CID 39337082.
- ↑ Journalist, Vibeke Lyngklip Svansø; Journalist, Signe Westermann Kühn (2023-06-23). "Ny bekymring om populære Novo Nordisk-lægemidler – undersøger om de kan give selvmordstanker". Berlingske.dk (in Danish). Retrieved 2023-07-04.
- ↑ Weber, Cordelia (2023-06-21). "Novo Nordisk-slankemiddel undersøges for at øge kræftrisiko". www.bt.dk (in Danish). Retrieved 2023-07-04.
- ↑ Fokos, S.; Panagis, G. (2010). "Effects of delta9-tetrahydrocannabinol on reward and anxiety in rats exposed to chronic unpredictable stress". Journal of Psychopharmacology. 24 (5): 767–777. doi:10.1177/0269881109104904. ISSN 1461-7285. PMID 19406854. S2CID 8855238.
- ↑ Sharpe, Lara; Sinclair, Justin; Kramer, Andrew; de Manincor, Michael; Sarris, Jerome (2020-10-02). "Cannabis, a cause for anxiety? A critical appraisal of the anxiogenic and anxiolytic properties". Journal of Translational Medicine. 18 (1): 374. doi:10.1186/s12967-020-02518-2. ISSN 1479-5876. PMC 7531079. PMID 33008420.
- ↑ Rafiei, Dorsa; Kolla, Nathan J. (2021-01-21). "Elevated Brain Fatty Acid Amide Hydrolase Induces Depressive-Like Phenotypes in Rodent Models: A Review". International Journal of Molecular Sciences. 22 (3): 1047. doi:10.3390/ijms22031047. ISSN 1422-0067. PMC 7864498. PMID 33494322.
- ↑ Viveros, M. P.; Marco, Eva M.; File, Sandra E. (2005). "Endocannabinoid system and stress and anxiety responses". Pharmacology, Biochemistry, and Behavior. 81 (2): 331–342. doi:10.1016/j.pbb.2005.01.029. ISSN 0091-3057. PMID 15927244. S2CID 25052842.
- ↑ Sipe JC, Scott TM, Murray S, Harismendy O, Simon GM, Cravatt BF, Waalen J (January 2010). "Biomarkers of endocannabinoid system activation in severe obesity". PLOS ONE. 5 (1): e8792. Bibcode:2010PLoSO...5.8792S. doi:10.1371/journal.pone.0008792. PMC 2808340. PMID 20098695.
- ↑ Balsevich G, Sticht M, Bowles NP, Singh A, Lee TT, Li Z, et al. (July 2018). "Role for fatty acid amide hydrolase (FAAH) in the leptin-mediated effects on feeding and energy balance". Proceedings of the National Academy of Sciences of the United States of America. 115 (29): 7605–7610. Bibcode:2018PNAS..115.7605B. doi:10.1073/pnas.1802251115. PMC 6055171. PMID 29967158.
- ↑ Trayhurn P, Bing C, Wood IS (July 2006). "Adipose tissue and adipokines--energy regulation from the human perspective". The Journal of Nutrition. 136 (7 Suppl): 1935S–1939S. doi:10.1093/jn/136.7.1935S. PMID 16772463.
- ↑ Cota D (2008). "Role of the endocannabinoid system in energy balance regulation and obesity". Obesity and Metabolism. Frontiers of Hormone Research. Vol. 36. Basel: KARGER. pp. 135–145. doi:10.1159/000115362. ISBN 978-3-8055-8429-6. PMID 18230900.
- ↑ Teixeira D, Pestana D, Faria A, Calhau C, Azevedo I, Monteiro R (November 2010). "Modulation of adipocyte biology by δ(9)-tetrahydrocannabinol". Obesity. 18 (11): 2077–2085. doi:10.1038/oby.2010.100. PMID 20467421.
- ↑ Pagano C, Pilon C, Calcagno A, Urbanet R, Rossato M, Milan G, et al. (December 2007). "The endogenous cannabinoid system stimulates glucose uptake in human fat cells via phosphatidylinositol 3-kinase and calcium-dependent mechanisms". The Journal of Clinical Endocrinology and Metabolism. 92 (12): 4810–4819. doi:10.1210/jc.2007-0768. PMID 17785353.
- ↑ Rajavashisth TB, Shaheen M, Norris KC, Pan D, Sinha SK, Ortega J, Friedman TC (2012-01-01). "Decreased prevalence of diabetes in marijuana users: cross-sectional data from the National Health and Nutrition Examination Survey (NHANES) III". BMJ Open. 2 (1): e000494. doi:10.1136/bmjopen-2011-000494. PMC 3289985. PMID 22368296.
- ↑ Clark TM, Jones JM, Hall AG, Tabner SA, Kmiec RL (2018-12-21). "Theoretical Explanation for Reduced Body Mass Index and Obesity Rates in Cannabis Users". Cannabis and Cannabinoid Research. 3 (1): 259–271. doi:10.1089/can.2018.0045. PMC 6340377. PMID 30671538.
- ↑ Danielsson AK, Lundin A, Yaregal A, Östenson CG, Allebeck P, Agardh EE (2016). "Cannabis Use as Risk or Protection for Type 2 Diabetes: A Longitudinal Study of 18 000 Swedish Men and Women". Journal of Diabetes Research. 2016: 6278709. doi:10.1155/2016/6278709. PMC 5098083. PMID 27843955.
- ↑ Farokhnia M, McDiarmid GR, Newmeyer MN, Munjal V, Abulseoud OA, Huestis MA, Leggio L (February 2020). "Effects of oral, smoked, and vaporized cannabis on endocrine pathways related to appetite and metabolism: a randomized, double-blind, placebo-controlled, human laboratory study". Translational Psychiatry. 10 (1): 71. doi:10.1038/s41398-020-0756-3. PMC 7031261. PMID 32075958.
- ↑ Adejumo AC, Alliu S, Ajayi TO, Adejumo KL, Adegbala OM, Onyeakusi NE, et al. (2017-04-25). "Cannabis use is associated with reduced prevalence of non-alcoholic fatty liver disease: A cross-sectional study". PLOS ONE. 12 (4): e0176416. Bibcode:2017PLoSO..1276416A. doi:10.1371/journal.pone.0176416. PMC 5404771. PMID 28441459.
- 1 2 Burstein SH (May 2014). "The cannabinoid acids, analogs and endogenous counterparts". Bioorganic & Medicinal Chemistry. 22 (10): 2830–2843. doi:10.1016/j.bmc.2014.03.038. PMC 4351512. PMID 24731541.
- ↑ Grotenhermen F (2003). "Pharmacokinetics and pharmacodynamics of cannabinoids". Clinical Pharmacokinetics. 42 (4): 327–360. doi:10.2165/00003088-200342040-00003. PMID 12648025. S2CID 25623600.
- ↑ Grotenhermen F (2003-04-01). "Pharmacokinetics and pharmacodynamics of cannabinoids". Clinical Pharmacokinetics. 42 (4): 327–360. doi:10.2165/00003088-200342040-00003. PMID 12648025. S2CID 25623600.
- ↑ Ujváry I, Grotenhermen F (2014). "11-Nor-9-carboxy-∆9-tetrahydrocannabinol – a ubiquitous yet under researched cannabinoid. A review of the literature" (PDF). cannabis-med.org. Archived from the original (PDF) on 2017-04-20. Retrieved 2021-04-05.
- ↑ Purschke K, Heinl S, Lerch O, Erdmann F, Veit F (June 2016). "Development and validation of an automated liquid-liquid extraction GC/MS method for the determination of THC, 11-OH-THC, and free THC-carboxylic acid (THC-COOH) from blood serum". Analytical and Bioanalytical Chemistry. 408 (16): 4379–4388. doi:10.1007/s00216-016-9537-5. PMC 4875941. PMID 27116418.
- 1 2 3 Nguyen BM, Kim D, Bricker S, Bongard F, Neville A, Putnam B, et al. (October 2014). "Effect of marijuana use on outcomes in traumatic brain injury". The American Surgeon. 80 (10): 979–983. doi:10.1177/000313481408001015. PMID 25264643. S2CID 25797119.
- 1 2 El-Gohary M, Eid MA (March 2004). "Effect of cannabinoid ingestion (in the form of bhang) on the immune system of high school and university students". Human & Experimental Toxicology. 23 (3): 149–156. Bibcode:2004HETox..23..149E. doi:10.1191/0960327104ht426oa. PMID 15119535. S2CID 23909791.
- ↑ Păunescu H, Coman OA, Coman L, Ghiţă I, Georgescu SR, Drăghia F, Fulga I (2011-02-15). "Cannabinoid system and cyclooxygenases inhibitors". Journal of Medicine and Life. 4 (1): 11–20. PMC 3056416. PMID 21505570.
- ↑ Russo EB (February 2008). "Cannabinoids in the management of difficult to treat pain". Therapeutics and Clinical Risk Management. 4 (1): 245–59. doi:10.2147/tcrm.s1928. PMC 2503660. PMID 18728714.
- ↑ Leweke FM, Piomelli D, Pahlisch F, Muhl D, Gerth CW, Hoyer C, et al. (March 2012). "Cannabidiol enhances anandamide signaling and alleviates psychotic symptoms of schizophrenia". Translational Psychiatry. 2 (3): e94. doi:10.1038/tp.2012.15. PMC 3316151. PMID 22832859.
- ↑ Mechoulam R (2019). "The Chemistry Behind Cannabinoid Acids (COOH)". YouTube. Archived from the original on 2021-12-15.
- 1 2 Kathuria S, Gaetani S, Fegley D, Valiño F, Duranti A, Tontini A, et al. (January 2003). "Modulation of anxiety through blockade of anandamide hydrolysis". Nature Medicine. 9 (1): 76–81. doi:10.1038/nm803. PMID 12461523. S2CID 20764484.
- ↑ Sanchez-Marin L, Pavon FJ, Decara J, Suarez J, Gavito A, Castilla-Ortega E, et al. (2017-02-07). "Effects of Intermittent Alcohol Exposure on Emotion and Cognition: A Potential Role for the Endogenous Cannabinoid System and Neuroinflammation". Frontiers in Behavioral Neuroscience. 11: 15. doi:10.3389/fnbeh.2017.00015. PMC 5293779. PMID 28223925.
- ↑ Mahmood OM, Jacobus J, Bava S, Scarlett A, Tapert SF (November 2010). "Learning and memory performances in adolescent users of alcohol and marijuana: interactive effects". Journal of Studies on Alcohol and Drugs. 71 (6): 885–894. doi:10.15288/jsad.2010.71.885. PMC 2965487. PMID 20946746.
- ↑ Schneider M, Kasanetz F, Lynch DL, Friemel CM, Lassalle O, Hurst DP, et al. (October 2015). "Enhanced Functional Activity of the Cannabinoid Type-1 Receptor Mediates Adolescent Behavior". The Journal of Neuroscience. 35 (41): 13975–13988. doi:10.1523/JNEUROSCI.1937-15.2015. PMC 4604232. PMID 26468198.
- ↑ Lutz B (February 2009). "Endocannabinoid signals in the control of emotion". Current Opinion in Pharmacology. Neurosciences. 9 (1): 46–52. doi:10.1016/j.coph.2008.12.001. PMID 19157983.
- ↑ Burns HD, Van Laere K, Sanabria-Bohórquez S, Hamill TG, Bormans G, Eng WS, et al. (June 2007). "[18F]MK-9470, a positron emission tomography (PET) tracer for in vivo human PET brain imaging of the cannabinoid-1 receptor". Proceedings of the National Academy of Sciences of the United States of America. 104 (23): 9800–9805. Bibcode:2007PNAS..104.9800B. doi:10.1073/pnas.0703472104. PMC 1877985. PMID 17535893.
- ↑ Grinspoon L (2009). "A Cannabis Odyssey: To Smoke or Not To Smoke by Lester Grinspoon". marijuana-uses.com.
- ↑ Garner, Emily (2016). "A phenomenology of marijuana use among graduate students". Indo-Pacific Journal of Phenomenology. 16 (1): 1–17. doi:10.1080/20797222.2016.1164997. ISSN 1445-7377. S2CID 148123749.
- ↑ Ferrer, Belen (2007). "Regulation of brain anandamide by acute administration of ethanol". The Biochemical Journal. 404 (Pt 1): 97–104. doi:10.1042/BJ20061898. PMC 1868843. PMID 17302558.
- ↑ PubChem. "Palmitoylethanolamide". pubchem.ncbi.nlm.nih.gov. Retrieved 2023-03-05.
- ↑ "N-arachidonoylphosphatidylethanolamine (CHEBI:52571)". www.ebi.ac.uk. Retrieved 2021-04-07.
- ↑ Hassanzadeh P, Arbabi E, Atyabi F, Dinarvand R (March 2016). "The endocannabinoid system and NGF are involved in the mechanism of action of resveratrol: a multi-target nutraceutical with therapeutic potential in neuropsychiatric disorders". Psychopharmacology. 233 (6): 1087–1096. doi:10.1007/s00213-015-4188-3. PMID 26780936. S2CID 15089814.
- ↑ Gertsch, Jürg; Leonti, Marco; Raduner, Stefan; Racz, Ildiko; Chen, Jian-Zhong; Xie, Xiang-Qun; Altmann, Karl-Heinz; Karsak, Meliha; Zimmer, Andreas (2008-07-01). "Beta-caryophyllene is a dietary cannabinoid". Proceedings of the National Academy of Sciences of the United States of America. 105 (26): 9099–9104. Bibcode:2008PNAS..105.9099G. doi:10.1073/pnas.0803601105. ISSN 0027-8424. PMC 2449371. PMID 18574142.
- ↑ Russo EB (July 2016). "Beyond Cannabis: Plants and the Endocannabinoid System". Trends in Pharmacological Sciences. 37 (7): 594–605. doi:10.1016/j.tips.2016.04.005. PMID 27179600.
- ↑ "Beyond Cannabis: Plants and the Endocannabinoid System". ResearchGate. Retrieved 2021-04-07.
- ↑ Scandiffio, Rosaria (2023-03-23). "Beta-Caryophyllene Modifies Intracellular Lipid Composition in a Cell Model of Hepatic Steatosis by Acting through CB2 and PPAR Receptors". Int J Mol Sci. 24 (7): 6060. doi:10.3390/ijms24076060. PMC 10094507. PMID 37047034.
- 1 2 Basavarajappa BS, Hungund BL (February 1999). "Chronic ethanol increases the cannabinoid receptor agonist anandamide and its precursor N-arachidonoylphosphatidylethanolamine in SK-N-SH cells". Journal of Neurochemistry. 72 (2): 522–528. doi:10.1046/j.1471-4159.1999.0720522.x. PMID 9930723. S2CID 10793743.
- ↑ Neumeister A, Normandin MD, Murrough JW, Henry S, Bailey CR, Luckenbaugh DA, et al. (December 2012). "Positron emission tomography shows elevated cannabinoid CB1 receptor binding in men with alcohol dependence". Alcoholism: Clinical and Experimental Research. 36 (12): 2104–2109. doi:10.1111/j.1530-0277.2012.01815.x. PMC 3418442. PMID 22551199.
- ↑ Hirvonen J, Zanotti-Fregonara P, Umhau JC, George DT, Rallis-Frutos D, Lyoo CH, et al. (August 2013). "Reduced cannabinoid CB1 receptor binding in alcohol dependence measured with positron emission tomography". Molecular Psychiatry. 18 (8): 916–921. doi:10.1038/mp.2012.100. PMC 3594469. PMID 22776901.
- ↑ Van Der Stelt M, Noordermeer MA, Kiss T, Van Zadelhoff G, Merghart B, Veldink GA, Vliegenthart JF (April 2000). "Formation of a new class of oxylipins from N-acyl(ethanol)amines by the lipoxygenase pathway". European Journal of Biochemistry. 267 (7): 2000–7. doi:10.1046/j.1432-1327.2000.01203.x. hdl:1874/5348. PMID 10727939. S2CID 42011572.
- ↑ Gachet MS, Schubert A, Calarco S, Boccard J, Gertsch J (January 2017). "Targeted metabolomics shows plasticity in the evolution of signaling lipids and uncovers old and new endocannabinoids in the plant kingdom". Scientific Reports. 7 (1): 41177. Bibcode:2017NatSR...741177G. doi:10.1038/srep41177. PMC 5264637. PMID 28120902.
- ↑ Chapman KD (November 2000). "Emerging physiological roles for N-acylphosphatidylethanolamine metabolism in plants: signal transduction and membrane protection". Chemistry and Physics of Lipids. 108 (1–2): 221–229. doi:10.1016/s0009-3084(00)00198-5. PMID 11106793.
- ↑ Castelli MP, Madeddu C, Casti A, Casu A, Casti P, Scherma M, et al. (2014-05-20). "Δ9-tetrahydrocannabinol prevents methamphetamine-induced neurotoxicity". PLOS ONE. 9 (5): e98079. Bibcode:2014PLoSO...998079C. doi:10.1371/journal.pone.0098079. PMC 4028295. PMID 24844285.
- ↑ Bregman T, Fride E (June 2011). "Treatment with tetrahydrocannabinol (THC) prevents infertility in male cystic fibrosis mice". Journal of Basic and Clinical Physiology and Pharmacology. 22 (1–2): 29–32. doi:10.1515/jbcpp.2011.004. PMID 22865360. S2CID 19335113.
- ↑ Russo E (2002). "Cannabis Treatments in Obstetrics and Gynecology: A Historical Review". cannabis-med.org.
- ↑ Bossong MG, van Hell HH, Jager G, Kahn RS, Ramsey NF, Jansma JM (December 2013). "The endocannabinoid system and emotional processing: a pharmacological fMRI study with ∆9-tetrahydrocannabinol". European Neuropsychopharmacology. 23 (12): 1687–1697. doi:10.1016/j.euroneuro.2013.06.009. PMID 23928295. S2CID 4847188.
- ↑ Sano K, Mishima K, Koushi E, Orito K, Egashira N, Irie K, et al. (January 2008). "Delta 9-tetrahydrocannabinol-induced catalepsy-like immobilization is mediated by decreased 5-HT neurotransmission in the nucleus accumbens due to the action of glutamate-containing neurons". Neuroscience. 151 (2): 320–328. doi:10.1016/j.neuroscience.2007.10.026. PMID 18083311. S2CID 45508434.
- ↑ Karhson DS, Hardan AY, Parker KJ (September 2016). "Endocannabinoid signaling in social functioning: an RDoC perspective". Translational Psychiatry. 6 (9): e905. doi:10.1038/tp.2016.169. PMC 5048207. PMID 27676446.
- 1 2 Hosking RD, Zajicek JP (July 2008). "Therapeutic potential of cannabis in pain medicine". British Journal of Anaesthesia. 101 (1): 59–68. doi:10.1093/bja/aen119. PMID 18515270.
- ↑ Howlett AC, Barth F, Bonner TI, Cabral G, Casellas P, Devane WA, et al. (June 2002). "International Union of Pharmacology. XXVII. Classification of cannabinoid receptors". Pharmacological Reviews. 54 (2): 161–202. doi:10.1124/pr.54.2.161. PMID 12037135. S2CID 8259002.
- ↑ Ripple W (2020). "World Scientists' Warning of a Climate Emergency". BioScience.
- ↑ Gillis J (2013-10-09). "By 2047, Coldest Years May Be Warmer Than Hottest in Past, Scientists Say". The New York Times. ISSN 0362-4331. Retrieved 2022-09-22.
- ↑ Mora C, Dousset B, Caldwell IR, Powell FE, Geronimo RC, Bielecki CR, et al. (2017-06-19). "Global risk of deadly heat". Nature Climate Change. 7 (7): 501–506. Bibcode:2017NatCC...7..501M. doi:10.1038/nclimate3322. ISSN 1758-678X. S2CID 90219036.
- ↑ Greenfield C, Dickie G (2022-06-14). "In hottest city on Earth, mothers bear brunt of climate change". Reuters. Retrieved 2022-10-21.
- ↑ Zaelke D (2008). "Reducing Black Carbon May Be the Fastest Strategy for Slowing Climate Change" (PDF). Institute for Governance & Sustainable Development.
- ↑ Meng L, Liu J, Tarasick DW, Randel WJ, Steiner AK, Wilhelmsen H, et al. (November 2021). "Continuous rise of the tropopause in the Northern Hemisphere over 1980-2020". Science Advances. 7 (45): eabi8065. Bibcode:2021SciA....7R8065M. doi:10.1126/sciadv.abi8065. PMC 8570593. PMID 34739322.
- ↑ "Warming temperatures increasingly alter structure of atmosphere | NCAR & UCAR News". news.ucar.edu. Retrieved 2022-09-22.
- ↑ Kim SY, Sung SA, Ko GJ, Boo CS, Jo SK, Cho WY, Kim HK (September 2006). "A case of multiple organ failure due to heat stoke following a warm bath". The Korean Journal of Internal Medicine. 21 (3): 210–212. doi:10.3904/kjim.2006.21.3.210. PMC 3890729. PMID 17017675.
- ↑ Rawls SM, Benamar K (June 2011). "Effects of opioids, cannabinoids, and vanilloids on body temperature". Frontiers in Bioscience. 3 (3): 822–845. doi:10.2741/190. PMC 3979758. PMID 21622235.
- ↑ Smirnov MS, Kiyatkin EA (September 2008). "Behavioral and temperature effects of delta 9-tetrahydrocannabinol in human-relevant doses in rats". Brain Research. 1228: 145–160. doi:10.1016/j.brainres.2008.06.069. PMC 2613277. PMID 18619424.
- ↑ Devane WA, Dysarz FA, Johnson MR, Melvin LS, Howlett AC (November 1988). "Determination and characterization of a cannabinoid receptor in rat brain". Molecular Pharmacology. 34 (5): 605–613. PMID 2848184.
- ↑ Gill, E. W.; Paton, W. D. M.; Pertwee, R. G. (1970-10-10). "Preliminary Experiments on the Chemistry and Pharmacology of Cannabis". Nature. 228 (5267): 134–136. Bibcode:1970Natur.228..134G. doi:10.1038/228134a0. ISSN 1476-4687. PMID 5466704. S2CID 4193316.
- ↑ Jung J, Meyer MR, Maurer HH, Neusüss C, Weinmann W, Auwärter V (October 2009). "Studies on the metabolism of the Delta9-tetrahydrocannabinol precursor Delta9-tetrahydrocannabinolic acid A (Delta9-THCA-A) in rat using LC-MS/MS, LC-QTOF MS and GC-MS techniques". Journal of Mass Spectrometry. 44 (10): 1423–1433. Bibcode:2009JMSp...44.1423J. doi:10.1002/jms.1624. PMID 19728318.
- ↑ Taghavi S, Ramirez S, Duchesne J, Tatum D (January 2021). "Preinjury Use of Marijuana and Outcomes in Trauma Patients". The Journal of Surgical Research. 257: 42–49. doi:10.1016/j.jss.2020.07.011. PMID 32818783. S2CID 221219765.
- ↑ Rieder SA, Chauhan A, Singh U, Nagarkatti M, Nagarkatti P (August 2010). "Cannabinoid-induced apoptosis in immune cells as a pathway to immunosuppression". Immunobiology. 215 (8): 598–605. doi:10.1016/j.imbio.2009.04.001. PMC 3005548. PMID 19457575.
- ↑ Hwang, Jeannie (2009-06-13). "Enhancement of endocannabinoid signaling by fatty acid amide hydrolase inhibition: A neuroprotective therapeutic modality". Life Sciences. 86 (15–16): 615–623. doi:10.1016/j.lfs.2009.06.003. PMC 2848893. PMID 19527737.
- ↑ Pollution Disease Link: Interview with neuroscience professor David Allsop from Lancaster University, retrieved 2022-10-26
- ↑ Eubanks LM, Rogers CJ, Beuscher AE, Koob GF, Olson AJ, Dickerson TJ, Janda KD (2006). "A molecular link between the active component of marijuana and Alzheimer's disease pathology". Molecular Pharmaceutics. 3 (6): 773–777. doi:10.1021/mp060066m. PMC 2562334. PMID 17140265.
- ↑ "Cannabinoids as antioxidants and neuroprotectants - Patent US-6630507-B1 - PubChem". pubchem.ncbi.nlm.nih.gov. Retrieved 2022-12-16.
- ↑ Aso, Ester; Ferrer, Isidre (2014-03-05). "Cannabinoids for treatment of Alzheimer's disease: moving toward the clinic". Frontiers in Pharmacology. 5: 37. doi:10.3389/fphar.2014.00037. ISSN 1663-9812. PMC 3942876. PMID 24634659.
- ↑ Coles, Madilyn; Steiner-Lim, Genevieve Z.; Karl, Tim (2022). "Therapeutic properties of multi-cannabinoid treatment strategies for Alzheimer's disease". Frontiers in Neuroscience. 16: 962922. doi:10.3389/fnins.2022.962922. ISSN 1662-453X. PMC 9479694. PMID 36117622.
- 1 2 "Cannabis Our Key to the Endocannabinoid System". www.caymanchem.com. Retrieved 2021-06-25.
- ↑ Steiner-Lim, Genevieve Z.; Coles, Madilyn; Jaye, Kayla; Metri, Najwa-Joelle; Butt, Ali S.; Christofides, Katerina; McPartland, Jackson; Al-Modhefer, Zainab; Karamacoska, Diana (2023). "Medicinal Cannabis for Alzheimer's Disease". ResearchGate. Retrieved 2023-08-13.
- ↑ Irenej, Pečan Luka; Francisco-Righoberto, Barrios; Marko, Jeran (2023). "Cannabinoid Molecules from Cannabis Sativa L. as a Promissing [sic] Solution for Methicillin-Resistant Staphylococcus Aureus (MRSA)" (PDF). University of Ljubljana.
- ↑ Farha, Maya A.; El-Halfawy, Omar M.; Gale, Robert T.; MacNair, Craig R.; Carfrae, Lindsey A.; Zhang, Xiong; Jentsch, Nicholas G.; Magolan, Jakob; Brown, Eric D. (2020-03-13). "Uncovering the Hidden Antibiotic Potential of Cannabis". ACS Infectious Diseases. 6 (3): 338–346. doi:10.1021/acsinfecdis.9b00419. ISSN 2373-8227. PMID 32017534. S2CID 211035245 – via ResearchGate.
- ↑ Rossi S, De Chiara V, Musella A, Mataluni G, Sacchetti L, Siracusano A, et al. (March 2009). "Caffeine drinking potentiates cannabinoid transmission in the striatum: interaction with stress effects". Neuropharmacology. 56 (3): 590–597. doi:10.1016/j.neuropharm.2008.10.013. PMID 19027757. S2CID 35191973.
- ↑ Rossi S, De Chiara V, Musella A, Mataluni G, Sacchetti L, Siracusano A, et al. (April 2010). "Effects of caffeine on striatal neurotransmission: focus on cannabinoid CB1 receptors". Molecular Nutrition & Food Research. 54 (4): 525–531. doi:10.1002/mnfr.200900237. PMID 20087854.
- ↑ Schelling G, Hauer D, Azad SC, Schmoelz M, Chouker A, Schmidt M, et al. (February 2006). "Effects of general anesthesia on anandamide blood levels in humans". Anesthesiology. 104 (2): 273–277. doi:10.1097/00000542-200602000-00012. PMID 16436846. S2CID 27303365.
- ↑ Patel S, Wohlfeil ER, Rademacher DJ, Carrier EJ, Perry LJ, Kundu A, et al. (July 2003). "The general anesthetic propofol increases brain N-arachidonylethanolamine (anandamide) content and inhibits fatty acid amide hydrolase". British Journal of Pharmacology. 139 (5): 1005–1013. doi:10.1038/sj.bjp.0705334. PMC 1573928. PMID 12839875.
- 1 2 de Melo Reis RA, Isaac AR, Freitas HR, de Almeida MM, Schuck PF, Ferreira GC, et al. (2021-10-28). "Quality of Life and a Surveillant Endocannabinoid System". Frontiers in Neuroscience. 15: 747229. doi:10.3389/fnins.2021.747229. PMC 8581450. PMID 34776851.
- ↑ Lauckner JE, Jensen JB, Chen HY, Lu HC, Hille B, Mackie K (February 2008). "GPR55 is a cannabinoid receptor that increases intracellular calcium and inhibits M current". Proceedings of the National Academy of Sciences of the United States of America. 105 (7): 2699–2704. Bibcode:2008PNAS..105.2699L. doi:10.1073/pnas.0711278105. PMC 2268199. PMID 18263732.
- ↑ Berk K, Bzdega W, Konstantynowicz-Nowicka K, Charytoniuk T, Zywno H, Chabowski A (January 2021). "Phytocannabinoids-A Green Approach toward Non-Alcoholic Fatty Liver Disease Treatment". Journal of Clinical Medicine. 10 (3): 393. doi:10.3390/jcm10030393. PMC 7864168. PMID 33498537.
- ↑ M Taalab Y, Mohammed WF, Helmy MA, Othman AA, Darwish M, Hassan I, Abbas M (November 2019). "Cannabis Influences the Putative Cytokines-Related Pathway of Epilepsy among Egyptian Epileptic Patients". Brain Sciences. 9 (12): 332. doi:10.3390/brainsci9120332. PMC 6955862. PMID 31757102.
- ↑ Hampson AJ, Grimaldi M, Axelrod J, Wink D (July 1998). "Cannabidiol and (-)Delta9-tetrahydrocannabinol are neuroprotective antioxidants". Proceedings of the National Academy of Sciences of the United States of America. 95 (14): 8268–8273. Bibcode:1998PNAS...95.8268H. doi:10.1073/pnas.95.14.8268. PMC 20965. PMID 9653176.
- ↑ D'Souza DC, Cortes-Briones JA, Ranganathan M, Thurnauer H, Creatura G, Surti T, et al. (January 2016). "Rapid Changes in CB1 Receptor Availability in Cannabis Dependent Males after Abstinence from Cannabis". Biological Psychiatry. Cognitive Neuroscience and Neuroimaging. 1 (1): 60–67. doi:10.1016/j.bpsc.2015.09.008. PMC 4742341. PMID 26858993.
- ↑ Ren M, Tang Z, Wu X, Spengler R, Jiang H, Yang Y, Boivin N (June 2019). "The origins of cannabis smoking: Chemical residue evidence from the first millennium BCE in the Pamirs". Science Advances. 5 (6): eaaw1391. Bibcode:2019SciA....5.1391R. doi:10.1126/sciadv.aaw1391. PMC 6561734. PMID 31206023.
- ↑ McPartland JM, Hegman W, Long (2019-11-01). "Cannabis in Asia: its center of origin and early cultivation, based on a synthesis of subfossil pollen and archaeobotanical studies". Vegetation History and Archaeobotany. 28 (6): 691–702. Bibcode:2019VegHA..28..691M. doi:10.1007/s00334-019-00731-8. ISSN 1617-6278. S2CID 181608199.
- ↑ Osterberger E, Lohwasser U, Jovanovic D, Ruzicka J, Novak J (2022-04-01). "The origin of the genus Cannabis". Genetic Resources and Crop Evolution. 69 (4): 1439–1449. doi:10.1007/s10722-021-01309-y. ISSN 1573-5109. S2CID 245589100.
- ↑ Korte G, Dreiseitel A, Schreier P, Oehme A, Locher S, Geiger S, et al. (January 2010). "Tea catechins' affinity for human cannabinoid receptors". Phytomedicine. 17 (1): 19–22. doi:10.1016/j.phymed.2009.10.001. PMID 19897346.
- ↑ Silver RJ (September 2019). "The Endocannabinoid System of Animals". Animals. 9 (9): 686. doi:10.3390/ani9090686. PMC 6770351. PMID 31527410.
- ↑ McPartland JM (2002). "Sourcing the Code: Searching for the Evolutionary Origins of Cannabinoid Receptors, Vanilloid Receptors, and Anandamide" (PDF). cannabis-med.org.
- ↑ Di Marzo, Vincenzo; De Petrocellis, Luciano (2012-12-05). "Why do cannabinoid receptors have more than one endogenous ligand?". Philosophical Transactions of the Royal Society B: Biological Sciences. 367 (1607): 3216–3228. doi:10.1098/rstb.2011.0382. ISSN 0962-8436. PMC 3481524. PMID 23108541.
- ↑ Mechoulam R (2019). "Research on Cannabinoids Over the Decades and What's to Come - Raphael Mechoulam". YouTube. Archived from the original on 2021-12-15.
- ↑ Bachhuber MA, Saloner B, Cunningham CO, Barry CL (October 2014). "Medical cannabis laws and opioid analgesic overdose mortality in the United States, 1999-2010". JAMA Internal Medicine. 174 (10): 1668–1673. doi:10.1001/jamainternmed.2014.4005. PMC 4392651. PMID 25154332.
- ↑ Bradford AC, Bradford WD, Abraham A, Bagwell Adams G (May 2018). "Association Between US State Medical Cannabis Laws and Opioid Prescribing in the Medicare Part D Population". JAMA Internal Medicine. 178 (5): 667–672. doi:10.1001/jamainternmed.2018.0266. PMC 6145794. PMID 29610897.
- ↑ Jeffries D (2006). "Jeffrey's Journey - Healing a Child's Violent Rages With Pot". YouTube. Archived from the original on 2021-12-15.
- ↑ Lucas P, Boyd S, Milloy MJ, Walsh Z (March 2021). "Cannabis Significantly Reduces the Use of Prescription Opioids and Improves Quality of Life in Authorized Patients: Results of a Large Prospective Study". Pain Medicine. 22 (3): 727–739. doi:10.1093/pm/pnaa396. PMC 7971472. PMID 33367882.
- ↑ Kruk-Slomka M, Dzik A, Budzynska B, Biala G (December 2017). "Endocannabinoid System: the Direct and Indirect Involvement in the Memory and Learning Processes-a Short Review". Molecular Neurobiology. 54 (10): 8332–8347. doi:10.1007/s12035-016-0313-5. PMC 5684264. PMID 27924524.
- ↑ Mechoulam R (2007). "Cannabinoid system in neuroprotection, and the importance of forgetting (9:08)". YouTube. Archived from the original on 2021-12-15.
- ↑ "Cannabinoids To the Neurons and Beyond". www.caymanchem.com. Retrieved 2021-04-11.
- ↑ Cox B (2011). "Star Death and the Creation of Elements - Wonders of the Universe: Stardust". YouTube. Archived from the original on 2021-12-15.
- ↑ Callahan MP (2011). "DNA Building Blocks Can Be Made in Space". YouTube. Archived from the original on 2021-12-15.
- ↑ "New Evidence That Comets Deposited Building Blocks of Life on Primordial Earth". www.newswise.com. Retrieved 2021-04-03.
- ↑ Schultz C (2011). "Amino Acids from Interstellar Space". YouTube. Archived from the original on 2021-12-15.
- ↑ Maccarrone M, Bab I, Bíró T, Cabral GA, Dey SK, Di Marzo V, et al. (May 2015). "Endocannabinoid signaling at the periphery: 50 years after THC". Trends in Pharmacological Sciences. 36 (5): 277–296. doi:10.1016/j.tips.2015.02.008. PMC 4420685. PMID 25796370.
- ↑ Zaleski DP, Seifert NA, Steber AL, Muckle MT, Loomis RA, Corby JF, et al. (2013-03-01). "Detection of E-Cyanomethanimine toward Sagittarius B2(N) in the Green Bank Telescope PRIMOS Survey". The Astrophysical Journal Letters. 765 (1): L10. arXiv:1302.0909. Bibcode:2013ApJ...765L..10Z. doi:10.1088/2041-8205/765/1/L10. ISSN 0004-637X. S2CID 53552345.
- ↑ Remijan T (2013). "DNA component found in interstellar space". YouTube. Archived from the original on 2021-12-15.
- ↑ "atom | lex.dk". Den Store Danske (in Danish). Retrieved 2022-12-18.
External links
- N-Acylphosphatidylethanolamines (NAPEs), N-acylethanolamines (NAEs) and Other Acylamides: Metabolism, Occurrence and Functions in Plants; Center for Plant Lipid Research, Department of Biological Sciences, University of North Texas, Denton, TX, USA