| Aquaporin | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 | |||||||||
| Identifiers | |||||||||
| Symbol | Aquaporin | ||||||||
| Pfam | PF00230 | ||||||||
| InterPro | IPR000425 | ||||||||
| PROSITE | PDOC00193 | ||||||||
| SCOP2 | 1fx8 / SCOPe / SUPFAM | ||||||||
| TCDB | 1.A.8 | ||||||||
| OPM superfamily | 7 | ||||||||
| OPM protein | 2zz9 | ||||||||
| |||||||||
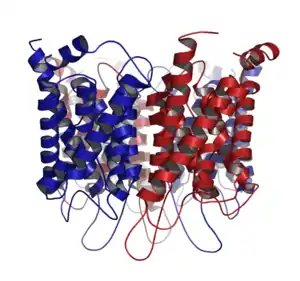
Aquaporins, also called water channels, are channel proteins from a larger family of major intrinsic proteins that form pores in the membrane of biological cells, mainly facilitating transport of water between cells.[1] The cell membranes of a variety of different bacteria, fungi, animal and plant cells contain aquaporins through which water can flow more rapidly into and out of the cell than by diffusing through the phospholipid bilayer.[2] Aquaporins have six membrane-spanning alpha helical domains with both carboxylic and amino terminals on the cytoplasmic side. Two hydrophobic loops contain conserved asparagine–proline–alanine ("NPA motif") which form a barrel surrounding a central pore-like region that contains additional protein density.[3] Because aquaporins are usually always open and are prevalent in just about every cell type, this leads to a misconception that water readily passes through the cell membrane down its concentration gradient. Water can pass through the cell membrane through simple diffusion because it is a small molecule, and through osmosis, in cases where the concentration of water outside of the cell is greater than that of the inside. However, because water is a polar molecule this process of simple diffusion is relatively slow, and in tissues with high water permeability the majority of water passes through aquaporin.[4][5]
The 2003 Nobel Prize in Chemistry was awarded jointly to Peter Agre for the discovery of aquaporins[6] and Roderick MacKinnon for his work on the structure and mechanism of potassium channels.[7]
Genetic defects involving aquaporin genes have been associated with several human diseases including nephrogenic diabetes insipidus and neuromyelitis optica.[8][9][10][11]
History
The mechanism of facilitated water transport and the probable existence of water pores has attracted researchers since 1957.[12] In most cells, water moves in and out by osmosis through the lipid component of cell membranes. Due to the relatively high water permeability of some epithelial cells, it was long suspected that some additional mechanism for water transport across membranes must exist. Solomon and his co-workers performed pioneering work on water permeability across the cell membrane in the late 1950s.[13][14] In the mid-1960s an alternative hypothesis (the "partition–diffusion model") sought to establish that the water molecules partitioned between the water phase and the lipid phase and then diffused through the membrane, crossing it until the next interphase where they left the lipid and returned to an aqueous phase.[15][16] Studies by Parisi, Edelman, Carvounis et al. accented not only the importance of the presence of water channels but also the possibility to regulate their permeability properties.[17][18][19] In 1990, Verkman's experiments demonstrated functional expression of water channels, indicating that water channels are effectively proteins.[20][21]
Discovery
It was not until 1992 that the first aquaporin, 'aquaporin-1' (originally known as CHIP 28), was reported by Peter Agre, of Johns Hopkins University.[22] In 1999, together with other research teams, Agre reported the first high-resolution images of the three-dimensional structure of an aquaporin, namely, aquaporin-1.[23] Further studies using supercomputer simulations identified the pathway of water as it moved through the channel and demonstrated how a pore can allow water to pass without the passage of small solutes.[24] The pioneering research and subsequent discovery of water channels by Agre and his colleagues won Agre the Nobel Prize in Chemistry in 2003.[7] Agre said he discovered aquaporins "by serendipity." He had been studying the Rh blood group antigens and had isolated the Rh molecule, but a second molecule, 28 kilodaltons in size (and therefore called 28K) kept appearing. At first they thought it was a Rh molecule fragment, or a contaminant, but it turned out to be a new kind of molecule with unknown function. It was present in structures such as kidney tubules and red blood cells, and related to proteins of diverse origins, such as in fruit fly brain, bacteria, the lens of the eye, and plant tissue.[23]
However the first report of protein-mediated water transport through membranes was by Gheorghe Benga and others in 1986, prior to Agre's first publication on the topic.[25][26] This led to a controversy that Benga's work had not been adequately recognized either by Agre or by the Nobel Prize Committee.[27]
Function
Aquaporins are "the plumbing system for cells". Water moves through cells in an organized way, most rapidly in tissues that have aquaporin water channels.[28] For many years, scientists assumed that water leaked through the cell membrane, and some water does. However, this did not explain how water could move so quickly through some cells.[28]
Aquaporins selectively conduct water molecules in and out of the cell, while preventing the passage of ions and other solutes. Also known as water channels, aquaporins are integral membrane pore proteins. Some of them, known as aquaglyceroporins, also transport other small uncharged dissolved molecules including ammonia, CO2, glycerol, and urea. For example, the aquaporin 3 channel has a pore width of 8–10 Ångströms and allows the passage of hydrophilic molecules ranging between 150 and 200 Da. However, the water pores completely block ions including protons, essential to conserve the membrane's electrochemical potential difference.[29]
Water molecules traverse through the pore of the channel in single file. The presence of water channels increases membrane permeability to water. These are also essential for the water transport system in plants[30] and tolerance to drought and salt stresses.[31]
Structure

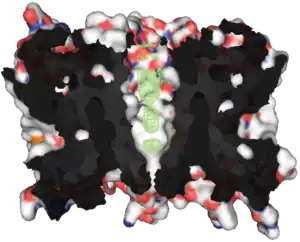
Aquaporin proteins are composed of a bundle of six transmembrane α-helices. They are embedded in the cell membrane. The amino and carboxyl ends face the inside of the cell. The amino and carboxyl halves resemble each other, apparently repeating a pattern of nucleotides. This may have been created by the doubling of a formerly half-sized gene. Between the helices are five regions (A – E) that loop into or out of the cell membrane, two of them hydrophobic (B, E), with an asparagine–proline–alanine ("NPA motif") pattern. They create a distinctive hourglass shape, making the water channel narrow in the middle and wider at each end.[29][32]
Another and even narrower place in the AQP1 channel is the "ar/R selectivity filter", a cluster of amino acids enabling the aquaporin to selectively let through or block the passage of different molecules.[33]
Aquaporins form four-part clusters (tetramers) in the cell membrane, with each of the four monomers acting as a water channel. Different aquaporins have different sized water channels, the smallest types allowing nothing but water through.[29]
X-ray profiles show that aquaporins have two conical entrances. This hourglass shape could be the result of a natural selection process toward optimal permeability.[34] It has been shown that conical entrances with suitable opening angle can indeed provide a large increase of the hydrodynamic channel permeability.[34]
NPA motif
Aquaporin channels appear in simulations to allow only water to pass, as the molecules effectively queue up in single file. Guided by the aquaporin's local electric field, the oxygen in each water molecule faces forwards as it enters, turning around half way along and leaving with the oxygen facing backwards.[35] The arrangement of opposite-facing electrostatic potentials in the two halves of the channel prevents the flow of protons but permits water to pass freely.[36]
ar/R selectivity filter
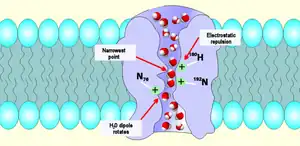
The aromatic/arginine or "ar/R" selectivity filter is a cluster of amino acids that help bind to water molecules and exclude other molecules that may try to enter the pore. It is the mechanism by which the aquaporin is able to selectively bind water molecules and so to allow them through, and to prevent other molecules from entering. The ar/R filter is made of two amino acid groups from helices B (HB) and E (HE) and two groups from loop E (LE1, LE2), from the two sides of the NPA motif. Its usual position is 8 Å on the outer side of the NPA motif; it is typically the tightest part of the channel. Its narrowness weakens the hydrogen bonds between water molecules, enabling the arginines, which carry a positive charge, to interact with the water molecules and to filter out undesirable protons.[37]
Taxonomic distribution
In mammals
There are thirteen known types of aquaporins in mammals; six of these are located in the kidney,[38] but the existence of many more is suspected. The most studied aquaporins are compared in the following table:
| Type | Location[39] | Function[39] |
|---|---|---|
| Aquaporin 1 | Water reabsorption | |
| Aquaporin 2 | Water reabsorption in response to ADH[40] | |
| Aquaporin 3 | Water reabsorption and glycerol permeability | |
| Aquaporin 4 | Water reabsorption |
In plants
In plants, water is taken up from the soil through the roots, where it passes from the cortex into the vascular tissues. There are three routes for water to flow in these tissues, known as the apoplastic, symplastic and transcellular pathways. Specifically, aquaporins are found in the vacuolar membrane, in addition to the plasma membrane of plants; the transcellular pathway involves transport of water across the plasma and vacuolar membranes.[41] When plant roots are exposed to mercuric chloride, which is known to inhibit aquaporins, the flow of water is greatly reduced while the flow of ions is not, supporting the view that there exists a mechanism for water transport independent of the transport of ions: aquaporins.[42] Aquaporins can play a major role in extension growth by allowing an influx of water into expanding cells - a process necessary to sustain plant development.[41] Plant aquaporins are important for mineral nutrition and ion detoxification; these are both essential for the homeostasis of minerals such as boron.[43]
Aquaporins in plants are separated into four main homologous subfamilies, or groups:[44]
- Plasma membrane Intrinsic Protein (PIP)[45]
- Tonoplast Intrinsic Protein (TIP)[46]
- Nodulin-26 like Intrinsic Protein (NIP)[47]
- Small basic Intrinsic Protein (SIP)[48]
These five subfamilies have later been divided into smaller evolutionary subgroups based on their DNA sequence. PIPs cluster into two subgroups, PIP1 and PIP2, whilst TIPs cluster into 5 subgroups, TIP1, TIP2, TIP3, TIP4 and TIP5. Each subgroup is again split up into isoforms e.g. PIP1;1, PIP1;2. As isoforms nomenclature are historically based on functional parameters rather than evolutive ones, several novel propositions on plant aquaporines have been arisen with the study of the evolutionary relationships between the different aquaporins.[49] Within the various selection of aquaporin isoforms in plants, there are also unique patterns of cell- and tissue-specific expression.[41]
When plant aquaporins are silenced, the hydraulic conductance and photosynthesis of the leaf decrease.[50] When gating of plant aquaporins occurs, it stops the flow of water through the pore of the protein. This may happen for various reasons, for example when the plant contains low amounts of cellular water due to drought.[51] The gating of an aquaporin is carried out by an interaction between a gating mechanism and the aquaporin, which causes a 3D change in the protein so that it blocks the pore and, thus, disallows the flow of water through the pore. In plants, there are at least two forms of aquaporin gating: gating by the dephosphorylation of certain serine residues, in response to drought, and the protonation of specific histidine residues, in response to flooding. The phosphorylation of an aquaporin is involved in the opening and closing of petals in response to temperature.[52][53]
In Heteroconts
Specific aquaporins called Large Intrinsic Proteins (LIP)[54] have been found in Heterokonts, including diatoms and brown algae. LIPs contain an NPM-motif in place of the second conserved NPA-motif typical of the majority of MIPs.
In other organisms
Aquaporins have been discovered in the fungi Saccharomyces cerevisiae (yeast), Dictyostelium, Candida and Ustilago and the protozoans Trypanosoma and Plasmodium.[30]
Clinical significance
There have been two clear examples of diseases identified as resulting from mutations in aquaporins: mutations in the aquaporin-2 gene cause hereditary nephrogenic diabetes insipidus in humans,[9] while mice homozygous for inactivating mutations in the aquaporin-0 gene develop congenital cataracts.[55] A small number of people have been identified with severe or total deficiency in aquaporin-1. They are, in general, healthy, but exhibit a defect in the ability to concentrate solutes in the urine and to conserve water when deprived of drinking water.[56][57] Mice with targeted deletions in aquaporin-1 also exhibit a deficiency in water conservation due to an inability to concentrate solutes in the kidney medulla by countercurrent multiplication.[58] Aquaporins play a key role in acquired forms of nephrogenic diabetes insipidus, disorders that cause increased urine production.[59] Aquaporin 2 is regulated by vasopressin which, when bound to the cell-surface receptor, activates the cAMP signaling pathway. This results in aquaporin-2 containing vesicles to increase water uptake and return to circulation. Mutation of the aquaporin 2 vasopressin receptor is a cause of acquired diabetes insipidus. In rats, acquired nephrogenic diabetes insipidus can be caused impaired regulation of aquaporin-2 due to administration of lithium salts, low potassium concentrations in the blood (hypokalemia) and high calcium concentrations in the blood (hypercalcemia).[60][61][62] Autoimmune reactions against aquaporin 4 in humans produce Devic's disease.[8] If aquaporin could be manipulated, that could potentially solve medical problems such as fluid retention in heart disease and brain edema after stroke.[28]
References
- ↑ Agre P (2006). "The aquaporin water channels". Proc Am Thorac Soc. 3 (1): 5–13. doi:10.1513/pats.200510-109JH. PMC 2658677. PMID 16493146.
- ↑ Cooper G (2009). The Cell: A Molecular Approach. Washington, DC: ASM PRESS. p. 544. ISBN 978-0-87893-300-6.
- ↑ Verkman, AS (January 2000). "Structure and function of aquaporin water channels". Am J Physiol Renal Physiol. 278 (1): F13-28. doi:10.1152/ajprenal.2000.278.1.F13. PMID 10644652.
- ↑ Cooper, Geoffrey (2000). The Cell (2 ed.). MA: Sinauer Associates. Retrieved 23 April 2020.
- ↑ Lodish, Harvey; Berk, Arnold; Zipursky, S. Lawrence (2000). Molecular Cell Biology (4th ed.). New York: W. H. Freeman. ISBN 9781464183393. Retrieved 20 May 2020.
- ↑ Knepper MA, Nielsen S (2004). "Peter Agre, 2003 Nobel Prize winner in chemistry". J. Am. Soc. Nephrol. 15 (4): 1093–5. doi:10.1097/01.ASN.0000118814.47663.7D. PMID 15034115.
- 1 2 "The Nobel Prize in Chemistry 2003". Nobel Foundation. Retrieved 2008-01-23.
- 1 2 Lennon VA, Kryzer TJ, Pittock SJ, Verkman AS, Hinson SR (2005). "IgG marker of optic-spinal multiple sclerosis binds to the aquaporin-4 water channel". J. Exp. Med. 202 (4): 473–7. doi:10.1084/jem.20050304. PMC 2212860. PMID 16087714.
- 1 2 Bichet DG (2006). "Nephrogenic diabetes insipidus" (PDF). Adv Chronic Kidney Dis. 13 (2): 96–104. doi:10.1053/j.ackd.2006.01.006. PMID 16580609. Archived from the original (PDF) on 2018-07-18.
- ↑ Agre P, Kozono D (2003). "Aquaporin water channels: molecular mechanisms for human diseases". FEBS Lett. 555 (1): 72–8. doi:10.1016/S0014-5793(03)01083-4. PMID 14630322. S2CID 35406097.
- ↑ Schrier RW (2007). "Aquaporin-related disorders of water homeostasis". Drug News Perspect. 20 (7): 447–53. doi:10.1358/dnp.2007.20.7.1138161. PMID 17992267.
- ↑ Parisi M, Dorr RA, Ozu M, Toriano R (December 2007). "From membrane pores to aquaporins: 50 years measuring water fluxes". J Biol Phys. 33 (5–6): 331–43. doi:10.1007/s10867-008-9064-5. PMC 2565768. PMID 19669522.
- ↑ Paganelli CV, Solomon AK (November 1957). "The rate of exchange of tritiated water across the human red cell membrane". J. Gen. Physiol. 41 (2): 259–77. doi:10.1085/jgp.41.2.259. PMC 2194835. PMID 13475690.
- ↑ Goldstein DA; Solomon AK (1960-09-01). "Determination of equivalent pore radius for human red cells by osmotic pressure measurement". The Journal of General Physiology. 44 (1): 1–17. doi:10.1085/jgp.44.1.1. PMC 2195086. PMID 13706631.
- ↑ Dainty, J.; House, C. R. (1966-07-01). "An examination of the evidence for membrane pores in frog skin". The Journal of Physiology. 185 (1): 172–184. doi:10.1113/jphysiol.1966.sp007979. PMC 1395865. PMID 5965891.
- ↑ Hanai T, Haydon DA (1966-08-01). "The permeability to water of bimolecular lipid membranes". Journal of Theoretical Biology. 11 (3): 370–382. Bibcode:1966JThBi..11..370H. doi:10.1016/0022-5193(66)90099-3. PMID 5967438.
- ↑ Parisi M, Bourguet J (1984-01-01). "Effects of cellular acidification on ADH-induced intramembrane particle aggregates". American Journal of Physiology. Cell Physiology. 246 (1): C157–C159. doi:10.1152/ajpcell.1984.246.1.c157. ISSN 0363-6143. PMID 6320654.
- ↑ Edelman, Isidore S. (25 May 1965). "Hydrogen-ion dependence of the antidiuretic action of vasopressin, oxytocin and deaminooxytocin". Biochimica et Biophysica Acta (BBA) - Biophysics Including Photosynthesis. 102 (1): 185–197. doi:10.1016/0926-6585(65)90212-8. PMID 5833400 – via Elsevier Science Direct.
- ↑ Carvounis CP, Levine SD, Hays RM (1979-05-01). "pH-Dependence of water and solute transport in toad urinary bladder". Kidney International. 15 (5): 513–519. doi:10.1038/ki.1979.66. ISSN 0085-2538. PMID 39188.
- ↑ Zhang, RB; Logee, KA; Verkman, AS (1990-09-15). "Expression of mRNA coding for kidney and red cell water channels in Xenopus oocytes". The Journal of Biological Chemistry. 265 (26): 15375–15378. doi:10.1016/S0021-9258(18)55405-3. ISSN 0021-9258. PMID 2394728.
- ↑ Zhang, R; Alper, S L; Thorens, B; Verkman, A S (1991-11-01). "Evidence from oocyte expression that the erythrocyte water channel is distinct from band 3 and the glucose transporter". Journal of Clinical Investigation. 88 (5): 1553–1558. doi:10.1172/JCI115466. PMC 295670. PMID 1939644.
- ↑ Agre P, Preston GM, Smith BL, Jung JS, Raina S, Moon C, Guggino WB, Nielsen S (1 October 1993). "Aquaporin CHIP: the archetypal molecular water channel". Am. J. Physiol. 265 (4 Pt 2): F463–76. doi:10.1152/ajprenal.1993.265.4.F463. PMID 7694481. S2CID 2685263.
- 1 2 Mitsuoka K, Murata K, Walz T, Hirai T, Agre P, Heymann JB, Engel A, Fujiyoshi Y (1999). "The structure of aquaporin-1 at 4.5-A resolution reveals short alpha-helices in the center of the monomer". J. Struct. Biol. 128 (1): 34–43. doi:10.1006/jsbi.1999.4177. PMID 10600556. S2CID 1076256.
- ↑ de Groot BL, Grubmüller H (2005). "The dynamics and energetics of water permeation and proton exclusion in aquaporins". Curr. Opin. Struct. Biol. 15 (2): 176–83. doi:10.1016/j.sbi.2005.02.003. hdl:11858/00-001M-0000-0012-E99D-E. PMID 15837176.
- ↑ Benga G, Popescu O, Pop VI, Holmes RP (1986). "p-(Chloromercuri)benzenesulfonate binding by membrane proteins and the inhibition of water transport in human erythrocytes". Biochemistry. 25 (7): 1535–8. doi:10.1021/bi00355a011. PMID 3011064.
- ↑ Kuchel PW (2006). "The story of the discovery of aquaporins: convergent evolution of ideas--but who got there first?". Cell. Mol. Biol. (Noisy-le-grand). 52 (7): 2–5. PMID 17543213.
- ↑ Benga, G. "Gheorghe Benga". Ad Astra - Online project for the Romanian Scientific Community. Archived from the original on December 25, 2007. Retrieved 2008-04-05.
- 1 2 3 A Conversation With Peter Agre: Using a Leadership Role to Put a Human Face on Science, By Claudia Dreifus, New York Times, January 26, 2009
- 1 2 3 Gonen T, Walz T (2006). "The structure of aquaporins". Q. Rev. Biophys. 39 (4): 361–96. doi:10.1017/S0033583506004458. PMID 17156589. S2CID 40235608.
- 1 2 Kruse E, Uehlein N, Kaldenhoff R (2006). "The aquaporins". Genome Biol. 7 (2): 206. doi:10.1186/gb-2006-7-2-206. PMC 1431727. PMID 16522221.
- ↑ Xu Y, et al. (2014). "A banana aquaporin gene". BMC Plant Biology. 14 (1): 59. doi:10.1186/1471-2229-14-59. PMC 4015420. PMID 24606771.
- ↑ Fu D, Lu M (2007). "The structural basis of water permeation and proton exclusion in aquaporins (Review)". Molecular Membrane Biology. 24 (5–6): 366–74. doi:10.1080/09687680701446965. PMID 17710641. S2CID 343461.
- ↑ Sui, Haixin; Han, Bong-Gyoon; Lee, John K.; Walian, Peter; Jap, Bing K. (2001). "Structural basis of water-specific transport through the AQP1 water channel". Nature. 414 (6866): 872–878. doi:10.1038/414872a. PMID 11780053.
- 1 2 Gravelle S, Joly L, Detcheverry F, Ybert C, Cottin-Bizonne C, Bocquet L (2013). "Optimizing water permeability through the hourglass shape of aquaporins". PNAS. 110 (41): 16367–16372. arXiv:1310.4309. Bibcode:2013PNAS..11016367G. doi:10.1073/pnas.1306447110. PMC 3799357. PMID 24067650.
- ↑ de Groot BL, Grubmüller H (2001). "Water permeation across biological membranes: mechanism and dynamics of aquaporin-1 and GlpF". Science. 294 (5550): 2353–2357. Bibcode:2001Sci...294.2353D. doi:10.1126/science.1062459. hdl:11858/00-001M-0000-0014-61AF-6. PMID 11743202. S2CID 446498.
- ↑ Tajkhorshid E, Nollert P, Jensen MØ, Miercke LJ, O'Connell J, Stroud RM, Schulten K (2002). "Control of the selectivity of the aquaporin water channel family by global orientational tuning". Science. 296 (5567): 525–30. Bibcode:2002Sci...296..525T. doi:10.1126/science.1067778. PMID 11964478. S2CID 22410850.
- ↑ Sui H, Han BG, Lee JK, Walian P, Jap BK (2001). "Structural basis of water-specific transport through the AQP1 water channel". Nature. 414 (6866): 872–878. doi:10.1038/414872a. PMID 11780053. S2CID 4315108.
- ↑ Nielsen S, Frøkiaer J, Marples D, Kwon TH, Agre P, Knepper MA (2002). "Aquaporins in the kidney: from molecules to medicine". Physiol. Rev. 82 (1): 205–44. doi:10.1152/physrev.00024.2001. PMID 11773613.
- 1 2 Unless else specified in table boxes, then ref is: Walter F. Boron (2005). Medical Physiology: A Cellular And Molecular Approaoch. Elsevier/Saunders. ISBN 978-1-4160-2328-9. Page 842
- ↑ Sands JM (2012). "Aquaporin 2: Not Just for Moving Water". Journal of the American Society of Nephrology. 23 (9): 1443–1444. doi:10.1681/ASN.2012060613. PMC 3431422. PMID 22797179.
- 1 2 3 Johansson, I; Karlsson, M; Johanson, U; Larsson, C; Kjellbom, P (2000-05-01). "The role of aquaporins in cellular and whole plant water balance". Biochimica et Biophysica Acta (BBA) - Biomembranes. 1465 (1–2): 324–342. doi:10.1016/S0005-2736(00)00147-4. PMID 10748263.
- ↑ Chaumont, F; Tyerman, SD (2014-04-01). "Aquaporins: Highly Regulated Channels Controlling Plant Water Relations". Plant Physiology. 164 (4): 1600–1618. doi:10.1104/pp.113.233791. PMC 3982727. PMID 24449709.
- ↑ Porcel, Rosa; Bustamante, Antonio; Ros, Roc; Serrano, Ramón; Mulet Salort, José M. (2018). "BvCOLD1: A novel aquaporin from sugar beet (Beta vulgaris L.) involved in boron homeostasis and abiotic stress". Plant, Cell & Environment. 41 (12): 2844–2857. doi:10.1111/pce.13416. hdl:10251/145984. PMID 30103284. S2CID 51974856.
- ↑ Kaldenhoff R, Bertl A, Otto B, Moshelion M, Uehlein N (2007). "Characterization of Plant Aquaporins". Osmosensing and Osmosignaling. Methods in Enzymology. Vol. 428. pp. 505–31. doi:10.1016/S0076-6879(07)28028-0. ISBN 978-0-12-373921-6. PMID 17875436.
{{cite book}}:|journal=ignored (help) - ↑ Kammerloher W, Fischer U, Piechottka GP, Schäffner AR (1994). "Water channels in the plant plasma membrane cloned by immunoselection from a mammalian expression system". Plant J. 6 (2): 187–99. doi:10.1046/j.1365-313X.1994.6020187.x. PMID 7920711.
- ↑ Maeshima M (2001). "TONOPLAST TRANSPORTERS: Organization and Function". Annu Rev Plant Physiol Plant Mol Biol. 52 (1): 469–497. doi:10.1146/annurev.arplant.52.1.469. PMID 11337406.
- ↑ Wallace IS, Choi WG, Roberts DM (2006). "The structure, function and regulation of the nodulin 26-like intrinsic protein family of plant aquaglyceroporins". Biochim. Biophys. Acta. 1758 (8): 1165–75. doi:10.1016/j.bbamem.2006.03.024. PMID 16716251.
- ↑ Johanson U, Gustavsson S (2002). "A new subfamily of major intrinsic proteins in plants". Mol. Biol. Evol. 19 (4): 456–61. doi:10.1093/oxfordjournals.molbev.a004101. PMID 11919287.
- ↑ Johanson, Urban; Karlsson, Maria; Johansson, Ingela; Gustavsson, Sofia; Sjövall, Sara; Fraysse, Laure; Weig, Alfons R.; Kjellbom, Per (2001). "The Complete Set of Genes Encoding Major Intrinsic Proteins in Arabidopsis Provides a Framework for a New Nomenclature for Major Intrinsic Proteins in Plants". Plant Physiology. 126 (4): 1358–1369. doi:10.1104/pp.126.4.1358. PMC 117137. PMID 11500536.
- ↑ Sade, N; Shatil-Cohen, A; Attia, Z; Maurel, C; Boursiac, Y; Kelly, G; Granot, D; Yaaran, A; Lerner, S (2014-11-01). "The Role of Plasma Membrane Aquaporins in Regulating the Bundle Sheath-Mesophyll Continuum and Leaf Hydraulics". Plant Physiology. 166 (3): 1609–1620. doi:10.1104/pp.114.248633. PMC 4226360. PMID 25266632.
- ↑ Kaldenhoff R, Fischer M (2006). "Aquaporins in plants". Acta Physiol (Oxf). 187 (1–2): 169–76. doi:10.1111/j.1748-1716.2006.01563.x. PMID 16734753. S2CID 35656554.
- ↑ Azad AK, Sawa Y, Ishikawa T, Shibata H (2004). "Phosphorylation of plasma membrane aquaporin regulates temperature-dependent opening of tulip petals". Plant Cell Physiol. 45 (5): 608–17. doi:10.1093/pcp/pch069. PMID 15169943.
- ↑ Azad AK, Katsuhara M, Sawa Y, Ishikawa T, Shibata H (2008). "Characterization of four plasma membrane aquaporins in tulip petals: a putative homolog is regulated by phosphorylation". Plant Cell Physiology. 49 (8): 1196–208. doi:10.1093/pcp/pcn095. PMID 18567892.
- ↑ Khabudaev KV, Petrova DP, Grachev MA, Likhoshway YV (2014). "A new subfamily LIP of the major intrinsic proteins". BMC Genomics. 15 (1): 1–7. doi:10.1186/1471-2164-15-173. PMC 4022174. PMID 24589353.
- ↑ Okamura T, Miyoshi I, Takahashi K, Mototani Y, Ishigaki S, Kon Y, Kasai N (2003). "Bilateral congenital cataracts result from a gain-of-function mutation in the gene for aquaporin-0 in mice". Genomics. 81 (4): 361–8. doi:10.1016/S0888-7543(03)00029-6. PMID 12676560.
- ↑ Radin, M. Judith; Yu, Ming-Jiun; Stoedkilde, Lene; Miller, R Lance; Hoffert, Jason D.; Frokiaer, Jorgen; Pisitkun, Trairak; Knepper, Mark A. (2017-03-06). "Aquaporin-2 Regulation in Health and Disease". Veterinary Clinical Pathology. 41 (4): 455–470. doi:10.1111/j.1939-165x.2012.00488.x. PMC 3562700. PMID 23130944.
- ↑ King, Landon S; Choi, Michael; Fernandez, Pedro C; Cartron, Jean-Pierre; Agre, Peter (2001-07-19). "Defective Urinary Concentrating Ability Due to a Complete Deficiency of Aquaporin-1". New England Journal of Medicine. 345 (3): 175–179. doi:10.1056/NEJM200107193450304. PMID 11463012.
- ↑ Schnermann, Jurgen; Chou, Chung-Lin; Ma, Tonghui; Traynor, Timothy; Knepper, Mark A; Verkman, AS (1998-08-04). "Defective proximal tubular fluid reabsorption in transgenic aquaporin-1 null mice". Proceedings of the National Academy of Sciences of the United States of America. 95 (16): 9660–9664. Bibcode:1998PNAS...95.9660S. doi:10.1073/pnas.95.16.9660. PMC 21395. PMID 9689137.
- ↑ Khanna A (2006). "Acquired nephrogenic diabetes insipidus". Semin. Nephrol. 26 (3): 244–8. doi:10.1016/j.semnephrol.2006.03.004. PMID 16713497.
- ↑ Christensen, S; Kusano, E; Yusufi, A N; Murayama, N; Dousa, TP (1985-06-01). "Pathogenesis of nephrogenic diabetes insipidus due to chronic administration of lithium in rats". Journal of Clinical Investigation. 75 (6): 1869–1879. doi:10.1172/JCI111901. PMC 425543. PMID 2989335.
- ↑ Marples, D; Frøkiaer, J; Dørup, J; Knepper, M A; Nielsen, S (1996-04-15). "Hypokalemia-induced downregulation of aquaporin-2 water channel expression in rat kidney medulla and cortex". Journal of Clinical Investigation. 97 (8): 1960–1968. doi:10.1172/JCI118628. PMC 507266. PMID 8621781.
- ↑ Marples, D; Christensen, S; Christensen, EI; Ottosen, P D; Nielsen, S (1995-04-01). "Lithium-induced downregulation of aquaporin-2 water channel expression in rat kidney medulla". Journal of Clinical Investigation. 95 (4): 1838–1845. doi:10.1172/JCI117863. PMC 295720. PMID 7535800.
External links
- Aquaporins at the U.S. National Library of Medicine Medical Subject Headings (MeSH)
- Animation (MPEG file at nobel.se)
- Computational Biomolecular Dynamics Group. "Aquaporin movies and pictures". Max Planck Institute. Archived from the original on April 25, 2006. Retrieved 2008-01-23.
- Theoretical and Computational Biophysics Group. "Structure, Dynamics, and Function of Aquaporins". University of Illinois at Urbana-Champaign. Retrieved 2008-01-23.