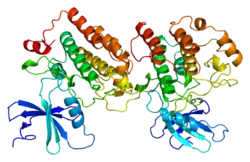



Casein kinase I isoform epsilon or CK1ε, is an enzyme that is encoded by the CSNK1E gene in humans.[5][6] It is the mammalian homolog of doubletime. CK1ε is a serine/threonine protein kinase and is very highly conserved; therefore, this kinase is very similar to other members of the casein kinase 1 family,[7] of which there are seven mammalian isoforms (α, β, γ1, γ2, γ3, δ and ε).[8] CK1ε is most similar to CK1δ in structure and function as the two enzymes maintain a high sequence similarity on their regulatory C-terminal and catalytic domains.[8] This gene is a major component of the mammalian oscillator which controls cellular circadian rhythms.[7] CK1ε has also been implicated in modulating various human health issues such as cancer, neurodegenerative diseases, and diabetes.[8]
Discovery
CK1ε-tau Mutation
In hamsters, the CK1ε-tau mutation was first discovered by Michael Menaker and Martin Ralph in 1988 while studying a laboratory shipment of Syrian hamsters.[9] They observed a hamster with an abnormal circadian period, and after breeding and further characterization, the two realized the mutation in hamsters conferred a shorter than normal free-running period.[9] They attributed this phenotype to what they termed the "tau mutation", which was the first full description of a mammalian circadian mutant.[10] This discovery provided a tool for other scientists to perform research on biological clocks and was an important early development in the field.[11]
Human CK1ε Cloned
In 1995, the human form of CK1ε was first isolated and cloned by the Virshup lab at the University of Utah.[12][13] It was officially identified as an isoform of the casein kinase 1 family.[12][13] Three transcript variants encoding the same protein have been found for this gene in rats: CK1ε1, CK1ε2, and CK1ε3; and two have been found in humans.[14][15]
Gene Mapping
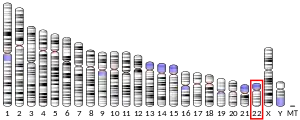
In 2000, the CK1ε gene was later mapped and identified by Joseph Takahashi and colleagues, who, using genetically directed representational difference analysis, discovered that the tau mutation was located on the CK1ε gene.[11] The CK1ε gene was found to be similar to the doubletime gene in Drosophila,[11] which had been first characterized and incorporated into biological clock function by Michael Young and colleagues in 1998.[16] In humans, the CSNK1E gene localizes at 22q13.1 and consists of 12 exons.[15]
Structural Imaging
Structural imaging was performed of CK1ε in 2012 by Alexander Long and colleagues using X-ray crystallography.[8] Certain kinase-related structural motifs were subsequently confirmed, such as a β-strand-turn-β-strand motif that anchors ATP, a DFG motif that orients ATP's phosphates, a catalytic loop that resembles that of PKA, and major substrate recognition sites in the C-terminal domain.[8]
Structure
The three-dimensional structures of the catalytic domains of the mammalian CK1δ and CK1ε were first solved by X-ray crystallography in 1996 and 2012 respectively.[8] CK1 kinase has multiple isoforms, including a total of seven characterized isoforms in mammals (alpha, beta, gamma1-3, delta, and epsilon.[17] The different isoforms differ mostly in the length and structure of their C-terminal non-catalytic region.[17] Only the delta and epsilon isoforms have been shown to play an important role in circadian rhythm regulation.[8]
CK1δ and CK1ε share a highly similar pattern in their structures.[17] The glycine rich P-loop is between the β1 and β2 strands, forming a classical β-strand-turn-β-strand motif that anchors and clamps the alpha phosphate of ATP.[8] CK1δ/ε additionally share conserved features within the catalytic domain, which are composed of both a N-terminal lobe and an α-helical C-terminal lobe.[8] The catalytic center is located in the cleft region between the two lobes, which also associates with the nucleotide and the substrate.[8] All known inhibitors bind to this center, blocking ATP binding.[17]
Function
Enzyme function
The protein encoded by the casein kinase 1 epsilon gene is a serine/threonine protein kinase and a member of the casein kinase I protein family, whose members have been implicated in the control of cytoplasmic and nuclear processes, including DNA replication and repair.[15] Like other casein kinase 1 protein family members, casein kinase 1 epsilon recognizes the Ser(p)XXSer/Thr motif for phosphorylation.[18] It is found in the cytoplasm as a monomer and can phosphorylate a variety of proteins, including itself.[15] This autophosphorylation occurs in the protein's C-Terminal domain, a region believed to behave as a pseudosubstrate, and inhibits kinase activity.[7][19][20]
The Circadian Clock
The Casein kinase 1 epsilon protein is part of the mammalian oscillator, a group of proteins that keep cells on a roughly 24-hour schedule.[21] This oscillator, or "circadian clock," is made up of a transcription-translation feedback loop (TTFL) in which several proteins work in tandem, each regulating the others' expression to generate a roughly 24-hour cycle of both mRNA and protein levels.[22] The TTFL also generates roughly 24-hour rhythms of outputs such as levels of cellular hormone release.[23] Daily oscillations in protein and mRNA transcription have been observed in many cells, including the mammalian master clock known as the suprachiasmatic nucleus (SCN).[24] However, unlike most circadian rhythm proteins that oscillate in their expression, casein kinase 1 epsilon is constitutively active.[23]
The core proteins that comprise the mammalian TTFL include Period (PER), Cryptochrome (CRY), BMAL1, CLOCK, and casein kinase 1 epsilon.[25] BMAL1 and CLOCK work to increase PER and CRY transcription by forming a heterodimer and binding on the E-box domain upstream from the PER and CRY gene coding sequences.[25] PER and CRY levels are regulated by negative feedback, meaning that they repress their own transcription.[25] Phosphorylation of PER proteins by CK1ε in both the cytoplasm and the nucleus marks these proteins for degradation.[26] Phosphorylation also hinders PER's ability to enter the nucleus by inducing a conformational change in its nuclear localization sequence.[7][27][28] On the other hand, the protein complex FBXL3 mediates the degradation of CRY proteins in the cytoplasm and nucleus.[29][30] If CRY binds to PER before it is phosphorylated by CK1ε, these three proteins stabilize into a complex that can enter the nucleus.[7] Once inside the nucleus, PER and CRY work to inhibit their own transcription while casein kinase 1 epsilon works to modulate the activity of BMAL1 and CLOCK through phosphorylation.[7]
As previously stated, the C-Terminal domain of casein kinase 1 epsilon behaves as a pseudosubstrate when phosphorylated, inhibiting kinase activity.[7][19][20] The C-Terminal domain has also been shown to be dephosphorylated by phosphatases such as Protein phosphatase 1 (PP1) in vitro and cell culture, which regulates levels of active casein kinase in vivo.[7][22][31] Current theory of circadian rhythms hypothesizes that this phosphorylation/dephosphorylation cycle of casein kinase 1 epsilon is important in modulation of the period of circadian rhythms in the cell, with increased phosphorylation decreasing casein kinase 1 epsilon activity (and subsequently increasing active CRY and PER) and dephosphorylation of casein kinase 1 epsilon resulting in a more active kinase (and lower levels of active CRY and PER).[22]
In mice, casein kinase 1 epsilon has been shown to phosphorylate both PER1 and PER2, as well as CRY1 and CRY2.[23] Casein kinase 1 results in a cyclic expression of mammalian oscillator proteins, resulting in a timekeeper (mammalian oscillator) for the cell:[32]
| Protein Level | Immediate Result | Delayed Result | |
|---|---|---|---|
| Dawn (7 am) | low PER and CRY protein concentration[33] | Per and Cry (gene) actively transcribed and stimulated by transcription factors BMAL1 and CLOCK | N/A |
| Dusk (7 pm) | high PER and CRY protein concentration[33] | high PER and CRY protein levels repress Per and Cry (gene) transcription | casein kinase 1 epsilon phosphorylates PER and CRY, marking the protein for degradation: PER and CRY protein concentration decreases |
Mutations to circadian function
The prominent phenotype in the CK1ε tau mutant hamsters discovered by Menaker was an unusually short free-running period — 22 hours in heterozygotes, and 20 hours in homozygotes for the mutation—making this allele semidominant.[34] The CK1ε gene was later mapped and identified by Joseph Takahashi and colleagues, which revealed a single base-pair C-to-T substitutional mutation in the hamster CK1ε gene.[35] This single nucleotide polymorphism (SNP) results in an arginine-to-cysteine substitution in a phosphate recognition domain region of CK1ε, a highly conserved region of the gene across mammals.[35] Presently, it is unclear how exactly the CK1ε-tau mutation results in a shorter free-running period.[36] However, it has been suggested that the tau mutation is a gain-of-function mutation, leading to increased phosphorylation of certain PER sites, thereby increasing the rate of PER degradation and shortening the circadian period.[37][21] The CK1ε-tau mutation in hamsters was the first full description of a mammalian circadian mutant.[10]
In humans, mutations affecting the PER2 phosphorylation site of the CK1ε and/or CK1δ gene result in Familial Advanced Sleep Phase Syndrome (FASPS).[38][39] This mutation, S662G, which results in the loss of a single phosphate acceptor site on PER2, prevents CK1ε protein from binding to PER and leads to an unusually short circadian period.[33]
Additionally, a heritable mutation in human CK1δ, T44A, has been identified as another mutation that causes period shortening, and it has been identified as another mechanism that causes FASPS.[40] This mutation reduces CK1δ activity in vivo in humans, and has similarly been shown to do the same in mice.[40] However, experiments in other species such as flies have shown that this mutation induces period lengthening effects.[40]
Furthermore, in humans, the P415A and H417R mutations in PER3 have been shown to destabilize the protein.[41] These mutations have been shown to generate FASPS and are also associated with impaired mood regulation.[41]
Temperature compensation
CK1δ/ε is temperature-compensated, a feature of many circadian rhythms.[42] The ability of CK1δ/ε to phosphorylate its substrates remains constant even when temperature fluctuates, whereas normal reactions rates tend to increase with increasing temperature.[42] Moreover, CK1ε tau mutants show a loss of temperature compensation.[42]
Non-mammalian homologs
Two circadian rhythm functional homologs of this mammalian protein can be found in Drosophila melanogaster (fruit fly).[43] Functional homologs refer to proteins sharing a similar function in another animal but that are not necessarily genetically similar.
One gene, coding for the protein Doubletime (abbreviated dbt), serves a similar purpose to casein kinase 1 epsilon in chronobiology, as it plays a role in the phosphorylation of PER.[7][43] However its gene sequence shows no sequence homology.[7][15][43][44] In addition, casein kinase 1 epsilon does not completely rescue circadian rhythms in fruit fly doubletime knockouts (dbt -/-), suggesting that these enzymes serve similar, but not identical, functions.[45][44]
Another functional homolog, the Drosophila gene for glycogen synthase kinase 3 (GSK3), called shaggy and abbreviated sgg, codes for a protein which phosphorylates Timeless (TIM), the fruit fly CRY functional homolog.[46] Like dbt, shaggy is not a sequence homolog to casein kinase 1 epsilon.[46] Conversely, Gsk3 is also found in mammals, and mutants have been implicated in circadian rhythm abnormalities in patients suffering with bipolar disorder.[7]
The Drosophila melanogaster genome contains other casein kinase 1 family enzymes, which are believed to serve no circadian function.[47] However, a different casein kinase family enzyme, casein kinase 2 alpha, has been implicated in providing the initial phosphorylation of a serine residue that is recognized by both DBT and Shaggy for sequential PER and TIM phosphorylation.[48][49]
Importance of CK1δ
While CK1ε has traditionally been considered the main regulator of PER and CRY phosphorylation, Casein kinase 1 isoform delta (CK1δ or CSNK1D), an isoform, is thought to play a similar role in the TTFL.[21] Both CK1ε and CK1δ phosphorylate and destabilize PER in vitro as well as interact with PER and CRY in vivo.[21] Moreover, CK1δ has been shown to better interact with proteins of the drosophila molecular clock than CK1ε, indicating that CK1δ may be more homologous to dbt than CK1ε.[21] Additionally, mass spectrometry has shown CK1δ to be over 20 times as abundant as CK1ε in the liver.[42]
Phosphoswitch Mechanism
Phosphorylation of PER2 is believed to be regulated by a phosphoswitch mechanism.[42] Specifically, PER2 requires an initial priming phosphorylation in order to be phosphorylated and subsequently degraded by CK1δ and/or CK1ε.[42] In this manner, temporally sequenced phosphorylations of PER2 act to delay its degradation rate and may provide insight into how the circadian clock is temperature compensated.[42] CK1δ and/or CK1ε may provide the priming activity.[42] The FASP site on PER2 is a key target of this priming kinase activity.[42] Mutations to this site can affect the ability of PER2 to receive a priming phosphorylation, leading to a lengthening or shortening of period.[42] Other studies have suggested that down stream phosphorylation of PER2 leads to stabilizing interactions that decrease the degradation rate of PER.[42] This is thought to increase the period of the circadian clock.[42] Mutations in the phosphorylation area of PER2 are thought to be related to FASPS patients[50]
Other functions
Canonical Wnt pathway
The canonical Wnt Pathway involves the accumulation of β-catenin in the cytoplasm, which activates transcription factors.[51] Casein kinase 1 epsilon, and related casein kinase 1 delta, is dephosphorylated in this pathway.[51][7] Dephosphorylation of casein kinase 1 epsilon is likely achieved by Protein Phosphatase 2 (PP2A), which increases both the enzymes' kinase activity in vivo.[7] Casein kinase 1 epsilon and casein kinase 1 delta have been implicated in increasing β-catenin's stability in the cytoplasm, although studies of the mechanism for this stabilization are inconclusive.[52] The current theory for how casein kinase 1 epsilon and/or casein kinase 1 delta function in this pathway is that both casein kinases either directly stabilize β-catenin though positive regulation, or that they indirectly stabilizes β-catenin through negative regulation of the β-catenin degradation complex (protease).[7][53]
Cancer
Casein kinase 1 epsilon and delta are known to phosphorylate a tumor suppressor protein, p53 in vivo in both humans and murine, or old world rats.[7][54][55][56] CK1 phosphorylates p53 on its N-terminus to induce its activation, which subsequently increases cell cycle arrest and apoptosis.[57] Damage to DNA has been shown to activate p53 through enhanced CK1 activation.[57] Inactivation of CK1 leads to decreased resistance to apoptosis.[57]
Casein kinase 1 epsilon is also implicated as indirectly causing cancer through its regulation of Yes-associated protein (YAP), an oncogene and regulator of organ size.[58] After priming through phosphorylation by the serine/threonine kinase LATS, both casein kinase 1 epsilon and casein kinase 1 delta have been shown to phosphorylate YAP and mark it for ubiquitination and degradation.[59]
Addiction
Several studies have demonstrated a connection between molecular components of the circadian clock and psychiatric disorders, particularly drug abuse.[60] Genetic association studies in humans have implicated CK1ε/CK1δ in the development of addictions to methamphetamine, heroin, and alcohol.[60] Moreover, mouse studies reveal a link between CK1ε/CK1δ activity and the stimulant effect produced by methamphetamine.[60] Additionally, inhibition of CK1ε/CK1δ in rodents has been shown to decrease alcohol and opiate relapse behavior during withdrawal.[61][62]
Interactions
Casein kinase 1 epsilon has been shown to interact with PER1,[28] PER2, CRY1, CRY2, BMAL1, CLOCK, NPAS2, and AXIN1.[7][63] PER1, PER2, and BMAL1 can be directly phosphorylated by CK1ɛ, while PER3, CRY1, and CRY2 can only be phosphorylated by CK1ɛ when associated with PER1 or PER2.[21]
Inhibitors
Several inhibitors have been produced by biotechnology companies to facilitate research on the function of casein kinase 1 epsilon. Testing utilizing CK1ε inhibitors have confirmed the involvement of CK1ε in a variety of processes, especially in regulation of circadian rhythms.
Pf-670462 and PF-4800567
PF-670462, developed by Pfizer, is a well-characterized inhibitor of both CK1ε and CK1δ that has been shown to lengthen the period of circadian rhythms when administered in vitro to rat fibroblasts and COS cells, and to mice in vivo.[21][64][65] PF-4800567, also developed by Pfizer, is a specific inhibitor of CK1ε. However, its ability to lengthen circadian rhythms is weaker than that of PF-670462 in both the in vitro rat fibroblasts and in vivo mice models.[65] The mechanisms of inhibition of PF-670462 and PF-4800567 differ between the two molecules as well.[8] PF-670462 maintains CK1ε/δ with the DFG motif facing inward, whereas PF-4800567 hydrophobically interacts with CK1ε/δ to turn the DFG motif outwards, indicative of a Type-II Kinase.[8]
IC261
IC261 is an inhibitor that targets the ATP binding site of both CK1δ and CK1ε.[21][66][57] Similarly, it has been shown to lengthen circadian period in rat fibroblasts,[66] and has been implicated in cancer treatment therapies for pancreatic and neuroblastomic cancers.[67][57]
Others
Other CK1 inhibitors, such as D4476, and pyrazolopyridine analogues, that both target CK1δ, have been characterized to have therapeutic abilities, but their beneficial effects are not well-studied and may stem from other cellular targets.[57]
See also
References
- 1 2 3 GRCh38: Ensembl release 89: ENSG00000213923 - Ensembl, May 2017
- 1 2 3 GRCm38: Ensembl release 89: ENSMUSG00000022433 - Ensembl, May 2017
- ↑ "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- ↑ "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- ↑ Fish KJ, Cegielska A, Getman ME, Landes GM, Virshup DM (June 1995). "Isolation and characterization of human casein kinase I epsilon (CKI), a novel member of the CKI gene family". The Journal of Biological Chemistry. 270 (25): 14875–83. doi:10.1074/jbc.270.25.14875. PMID 7797465.
- ↑ Sakanaka C, Leong P, Xu L, Harrison SD, Williams LT (October 1999). "Casein kinase iepsilon in the wnt pathway: regulation of beta-catenin function". Proceedings of the National Academy of Sciences of the United States of America. 96 (22): 12548–52. doi:10.1073/pnas.96.22.12548. PMC 22983. PMID 10535959.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 Knippschild U, Gocht A, Wolff S, Huber N, Löhler J, Stöter M (June 2005). "The casein kinase 1 family: participation in multiple cellular processes in eukaryotes". Cellular Signalling. 17 (6): 675–89. doi:10.1016/j.cellsig.2004.12.011. PMID 15722192.
- 1 2 3 4 5 6 7 8 9 10 11 12 Yang Y, Xu T, Zhang Y, Qin X (February 2017). "Molecular basis for the regulation of the circadian clock kinases CK1δ and CK1ε". Cellular Signalling. 31: 58–65. doi:10.1016/j.cellsig.2016.12.010. PMID 28057520.
- 1 2 Rensberger B (12 September 1988). "'Clock gene' may set 20-hour daily cycle for mutant hamsters". Washington Post.
- 1 2 Golombek DA, Rosenstein RE (July 2010). "Physiology of circadian entrainment". Physiological Reviews. 90 (3): 1063–102. doi:10.1152/physrev.00009.2009. hdl:11336/17613. PMID 20664079.
- 1 2 3 Lowrey PL, Shimomura K, Antoch MP, Yamazaki S, Zemenides PD, Ralph MR, Menaker M, Takahashi JS (April 2000). "Positional syntenic cloning and functional characterization of the mammalian circadian mutation tau". Science. 288 (5465): 483–92. Bibcode:2000Sci...288..483L. doi:10.1126/science.288.5465.483. PMC 3869379. PMID 10775102.
- "Gene Mutation Upsets Mammalian Biological Clock". Howard Hughes Medical Institute. April 21, 2000.
- 1 2 Fish KJ, Cegielska A, Getman ME, Landes GM, Virshup DM (June 1995). "Isolation and characterization of human casein kinase I epsilon (CKI), a novel member of the CKI gene family". The Journal of Biological Chemistry. 270 (25): 14875–83. doi:10.1074/jbc.270.25.14875. PMID 7797465.
- 1 2 Rodriguez N, Yang J, Hasselblatt K, Liu S, Zhou Y, Rauh-Hain JA, et al. (September 2012). "Casein kinase I epsilon interacts with mitochondrial proteins for the growth and survival of human ovarian cancer cells". EMBO Molecular Medicine. 4 (9): 952–63. doi:10.1002/emmm.201101094. PMC 3491827. PMID 22707389.
- ↑ Albrecht U (2010-01-23). The Circadian Clock. Springer Science & Business Media. ISBN 9781441912626.
- 1 2 3 4 5 "Entrez Gene: CSNK1E casein kinase 1, epsilon".
- ↑ Price JL, Blau J, Rothenfluh A, Abodeely M, Kloss B, Young MW (July 1998). "double-time is a novel Drosophila clock gene that regulates PERIOD protein accumulation". Cell. 94 (1): 83–95. doi:10.1016/S0092-8674(00)81224-6. PMID 9674430.
- 1 2 3 4 "NCBI CDD CDD Conserved Protein Domain STKc_CK1_delta_epsilon". www.ncbi.nlm.nih.gov. Retrieved 2019-04-11.
- ↑ Niefind K, Guerra B, Pinna LA, Issinger OG, Schomburg D (May 1998). "Crystal structure of the catalytic subunit of protein kinase CK2 from Zea mays at 2.1 A resolution". The EMBO Journal. 17 (9): 2451–62. doi:10.1093/emboj/17.9.2451. PMC 1170587. PMID 9564028.
- 1 2 Graves PR, Roach PJ (September 1995). "Role of COOH-terminal phosphorylation in the regulation of casein kinase I delta". The Journal of Biological Chemistry. 270 (37): 21689–94. doi:10.1074/jbc.270.37.21689. PMID 7665585.
- 1 2 Klimczak LJ, Farini D, Lin C, Ponti D, Cashmore AR, Giuliano G (October 1995). "Multiple isoforms of Arabidopsis casein kinase I combine conserved catalytic domains with variable carboxyl-terminal extensions". Plant Physiology. 109 (2): 687–96. doi:10.1104/pp.109.2.687. PMC 157637. PMID 7480353.
- 1 2 3 4 5 6 7 8 Etchegaray JP, Machida KK, Noton E, Constance CM, Dallmann R, Di Napoli MN, DeBruyne JP, Lambert CM, Yu EA, Reppert SM, Weaver DR (July 2009). "Casein kinase 1 delta regulates the pace of the mammalian circadian clock". Molecular and Cellular Biology. 29 (14): 3853–66. doi:10.1128/MCB.00338-09. PMC 2704743. PMID 19414593.
- 1 2 3 Richards J, Gumz ML (September 2012). "Advances in understanding the peripheral circadian clocks". FASEB Journal. 26 (9): 3602–13. doi:10.1096/fj.12-203554. PMC 3425819. PMID 22661008.
- 1 2 3 Ko CH, Takahashi JS (October 2006). "Molecular components of the mammalian circadian clock". Human Molecular Genetics. 15 Spec No 2 (suppl 2): R271-7. doi:10.1093/hmg/ddl207. PMC 3762864. PMID 16987893.
- ↑ Meijer JH, Michel S, Vanderleest HT, Rohling JH (December 2010). "Daily and seasonal adaptation of the circadian clock requires plasticity of the SCN neuronal network". The European Journal of Neuroscience. 32 (12): 2143–51. doi:10.1111/j.1460-9568.2010.07522.x. PMID 21143668. S2CID 12754517.
- 1 2 3 Takahashi JS (March 2017). "Transcriptional architecture of the mammalian circadian clock". Nature Reviews Genetics. 18 (3): 164–179. doi:10.1038/nrg.2016.150. PMC 5501165. PMID 27990019.
- ↑ Blau J (July 2008). "PERspective on PER phosphorylation". Genes & Development. 22 (13): 1737–40. doi:10.1101/gad.1696408. PMC 2732424. PMID 18593875.
- ↑ Akashi M, Tsuchiya Y, Yoshino T, Nishida E (March 2002). "Control of intracellular dynamics of mammalian period proteins by casein kinase I epsilon (CKIepsilon) and CKIdelta in cultured cells". Molecular and Cellular Biology. 22 (6): 1693–703. doi:10.1128/MCB.22.6.1693-1703.2002. PMC 135601. PMID 11865049.
- 1 2 Vielhaber E, Eide E, Rivers A, Gao ZH, Virshup DM (July 2000). "Nuclear entry of the circadian regulator mPER1 is controlled by mammalian casein kinase I epsilon". Molecular and Cellular Biology. 20 (13): 4888–99. doi:10.1128/MCB.20.13.4888-4899.2000. PMC 85940. PMID 10848614.
- ↑ Liu N, Zhang EE (2016-09-23). "Phosphorylation Regulating the Ratio of Intracellular CRY1 Protein Determines the Circadian Period". Frontiers in Neurology. 7: 159. doi:10.3389/fneur.2016.00159. PMC 5033960. PMID 27721804.
- ↑ Yoo SH, Mohawk JA, Siepka SM, Shan Y, Huh SK, Hong HK, Kornblum I, Kumar V, Koike N, Xu M, Nussbaum J, Liu X, Chen Z, Chen ZJ, Green CB, Takahashi JS (February 2013). "Competing E3 ubiquitin ligases govern circadian periodicity by degradation of CRY in nucleus and cytoplasm". Cell. 152 (5): 1091–105. doi:10.1016/j.cell.2013.01.055. PMC 3694781. PMID 23452855.
- ↑ Gietzen KF, Virshup DM (November 1999). "Identification of inhibitory autophosphorylation sites in casein kinase I epsilon". The Journal of Biological Chemistry. 274 (45): 32063–70. doi:10.1074/jbc.274.45.32063. PMID 10542239.
- ↑ Eide EJ, Kang H, Crapo S, Gallego M, Virshup DM (2005). "Casein kinase I in the mammalian circadian clock". Circadian Rhythms. Methods in Enzymology. Vol. 393. pp. 408–18. doi:10.1016/S0076-6879(05)93019-X. ISBN 9780121827984. PMC 1513158. PMID 15817302.
- 1 2 3 Partch CL, Green CB, Takahashi JS (February 2014). "Molecular architecture of the mammalian circadian clock". Trends in Cell Biology. 24 (2): 90–9. doi:10.1016/j.tcb.2013.07.002. PMC 3946763. PMID 23916625.
- ↑ Xiong Y, Zhou L, Su Z, Song J, Sun Q, Liu SS, Xia Y, Wang Z, Lu D (2019-02-05). "Longdaysin inhibits Wnt/β-catenin signaling and exhibits antitumor activity against breast cancer". OncoTargets and Therapy. 12: 993–1005. doi:10.2147/OTT.S193024. PMC 6368421. PMID 30787621.
- 1 2 Loudon AS, Meng QJ, Maywood ES, Bechtold DA, Boot-Handford RP, Hastings MH (2007). "The biology of the circadian Ck1epsilon tau mutation in mice and Syrian hamsters: a tale of two species". Cold Spring Harbor Symposia on Quantitative Biology. 72: 261–71. doi:10.1101/sqb.2007.72.073. PMID 18522517.
- ↑ Meng QJ, Logunova L, Maywood ES, Gallego M, Lebiecki J, Brown TM, Sládek M, Semikhodskii AS, Glossop NR, Piggins HD, Chesham JE, Bechtold DA, Yoo SH, Takahashi JS, Virshup DM, Boot-Handford RP, Hastings MH, Loudon AS (April 2008). "Setting clock speed in mammals: the CK1 epsilon tau mutation in mice accelerates circadian pacemakers by selectively destabilizing PERIOD proteins". Neuron. 58 (1): 78–88. doi:10.1016/j.neuron.2008.01.019. PMC 3756141. PMID 18400165.
- ↑ Eng GW, Virshup DM (2017-05-17). "Site-specific phosphorylation of casein kinase 1 δ (CK1δ) regulates its activity towards the circadian regulator PER2". PLOS ONE. 12 (5): e0177834. Bibcode:2017PLoSO..1277834E. doi:10.1371/journal.pone.0177834. PMC 5435336. PMID 28545154.
- ↑ Xu Y, Padiath QS, Shapiro RE, Jones CR, Wu SC, Saigoh N, Saigoh K, Ptácek LJ, Fu YH (March 2005). "Functional consequences of a CKIdelta mutation causing familial advanced sleep phase syndrome". Nature. 434 (7033): 640–4. Bibcode:2005Natur.434..640X. doi:10.1038/nature03453. PMID 15800623. S2CID 4416575.
- ↑ Fustin JM, Kojima R, Itoh K, Chang HY, Ye S, Zhuang B, Oji A, Gibo S, Narasimamurthy R, Virshup D, Kurosawa G, Doi M, Manabe I, Ishihama Y, Ikawa M, Okamura H (June 2018). "Ck1δ transcripts regulated by m6A methylation code for two antagonistic kinases in the control of the circadian clock". Proceedings of the National Academy of Sciences of the United States of America. 115 (23): 5980–5985. Bibcode:2018PNAS..115.5980F. doi:10.1073/pnas.1721371115. PMC 6003373. PMID 29784786.
- 1 2 3 Mignot E, Takahashi JS (January 2007). "A circadian sleep disorder reveals a complex clock". Cell. 128 (1): 22–3. doi:10.1016/j.cell.2006.12.024. PMC 3758472. PMID 17218251.
- 1 2 Melo MC, Abreu RL, Linhares Neto VB, de Bruin PF, de Bruin VM (August 2017). "Chronotype and circadian rhythm in bipolar disorder: A systematic review". Sleep Medicine Reviews. 34: 46–58. doi:10.1016/j.smrv.2016.06.007. PMID 27524206.
- 1 2 3 4 5 6 7 8 9 10 11 12 Narasimamurthy R, Hunt SR, Lu Y, Fustin JM, Okamura H, Partch CL, Forger DB, Kim JK, Virshup DM (June 2018). "CK1δ/ε protein kinase primes the PER2 circadian phosphoswitch". Proceedings of the National Academy of Sciences of the United States of America. 115 (23): 5986–5991. Bibcode:2018PNAS..115.5986N. doi:10.1073/pnas.1721076115. PMC 6003379. PMID 29784789.
- 1 2 3 Yu W, Zheng H, Price JL, Hardin PE (March 2009). "DOUBLETIME plays a noncatalytic role to mediate CLOCK phosphorylation and repress CLOCK-dependent transcription within the Drosophila circadian clock". Molecular and Cellular Biology. 29 (6): 1452–8. doi:10.1128/MCB.01777-08. PMC 2648245. PMID 19139270.
- 1 2 Lee H, Chen R, Lee Y, Yoo S, Lee C (December 2009). "Essential roles of CKIdelta and CKIepsilon in the mammalian circadian clock". Proceedings of the National Academy of Sciences of the United States of America. 106 (50): 21359–64. doi:10.1073/pnas.0906651106. PMC 2795500. PMID 19948962.
- ↑ Fan JY, Agyekum B, Venkatesan A, Hall DR, Keightley A, Bjes ES, Bouyain S, Price JL (November 2013). "Noncanonical FK506-binding protein BDBT binds DBT to enhance its circadian function and forms foci at night". Neuron. 80 (4): 984–96. doi:10.1016/j.neuron.2013.08.004. PMC 3869642. PMID 24210908.
- 1 2 Harms E, Young MW, Saez L (2003). "CK1 and GSK3 in the Drosophila and Mammalian Circadian Clock". Molecular Clocks and Light Signalling. Novartis Foundation Symposium. Vol. 253. pp. 267–77, discussion 102–9, 277–84. doi:10.1002/0470090839.ch19. ISBN 9780470852835. PMID 14712927.
- ↑ Marin O, Bustos VH, Cesaro L, Meggio F, Pagano MA, Antonelli M, Allende CC, Pinna LA, Allende JE (September 2003). "A noncanonical sequence phosphorylated by casein kinase 1 in beta-catenin may play a role in casein kinase 1 targeting of important signaling proteins". Proceedings of the National Academy of Sciences of the United States of America. 100 (18): 10193–200. Bibcode:2003PNAS..10010193M. doi:10.1073/pnas.1733909100. PMC 193538. PMID 12925738.
- ↑ Knippschild U, Gocht A, Wolff S, Huber N, Löhler J, Stöter M (June 2005). "The casein kinase 1 family: participation in multiple cellular processes in eukaryotes". Cellular Signalling. 17 (6): 675–89. doi:10.1016/j.cellsig.2004.12.011. PMID 15722192.
- ↑ Chiu JC, Vanselow JT, Kramer A, Edery I (July 2008). "The phospho-occupancy of an atypical SLIMB-binding site on PERIOD that is phosphorylated by DOUBLETIME controls the pace of the clock". Genes & Development. 22 (13): 1758–72. doi:10.1101/gad.1682708. PMC 2492663. PMID 18593878.
- ↑ Eng GW, Virshup DM (2017). "Site-specific phosphorylation of casein kinase 1 δ (CK1δ) regulates its activity towards the circadian regulator PER2". PLOS ONE. 12 (5): e0177834. Bibcode:2017PLoSO..1277834E. doi:10.1371/journal.pone.0177834. PMC 5435336. PMID 28545154.
- 1 2 Minde DP, Anvarian Z, Rüdiger SG, Maurice MM (August 2011). "Messing up disorder: how do missense mutations in the tumor suppressor protein APC lead to cancer?". Molecular Cancer. 10: 101. doi:10.1186/1476-4598-10-101. PMC 3170638. PMID 21859464.
- ↑ Lin SH, Lin YM, Yeh CM, Chen CJ, Chen MW, Hung HF, Yeh KT, Yang SF (February 2014). "Casein kinase 1 epsilon expression predicts poorer prognosis in low T-stage oral cancer patients". International Journal of Molecular Sciences. 15 (2): 2876–91. doi:10.3390/ijms15022876. PMC 3958887. PMID 24557581.
- ↑ Gao ZH, Seeling JM, Hill V, Yochum A, Virshup DM (February 2002). "Casein kinase I phosphorylates and destabilizes the beta-catenin degradation complex". Proceedings of the National Academy of Sciences of the United States of America. 99 (3): 1182–7. Bibcode:2002PNAS...99.1182G. doi:10.1073/pnas.032468199. PMC 122164. PMID 11818547.
- ↑ Knippschild U, Milne DM, Campbell LE, DeMaggio AJ, Christenson E, Hoekstra MF, Meek DW (October 1997). "p53 is phosphorylated in vitro and in vivo by the delta and epsilon isoforms of casein kinase 1 and enhances the level of casein kinase 1 delta in response to topoisomerase-directed drugs". Oncogene. 15 (14): 1727–36. doi:10.1038/sj.onc.1201541. PMID 9349507. S2CID 13476342.
- ↑ Shieh SY, Ikeda M, Taya Y, Prives C (October 1997). "DNA damage-induced phosphorylation of p53 alleviates inhibition by MDM2". Cell. 91 (3): 325–34. doi:10.1016/s0092-8674(00)80416-x. PMID 9363941.
- ↑ Sakaguchi K, Saito S, Higashimoto Y, Roy S, Anderson CW, Appella E (March 2000). "Damage-mediated phosphorylation of human p53 threonine 18 through a cascade mediated by a casein 1-like kinase. Effect on Mdm2 binding". The Journal of Biological Chemistry. 275 (13): 9278–83. doi:10.1074/jbc.275.13.9278. PMID 10734067.
- 1 2 3 4 5 6 Schittek B, Sinnberg T (October 2014). "Biological functions of casein kinase 1 isoforms and putative roles in tumorigenesis". Molecular Cancer. 13 (1): 231. doi:10.1186/1476-4598-13-231. PMC 4201705. PMID 25306547.
- ↑ Yu FX, Zhao B, Guan KL (November 2015). "Hippo Pathway in Organ Size Control, Tissue Homeostasis, and Cancer". Cell. 163 (4): 811–28. doi:10.1016/j.cell.2015.10.044. PMC 4638384. PMID 26544935.
- ↑ Zhou Q, Li L, Zhao B, Guan KL (April 2015). "The hippo pathway in heart development, regeneration, and diseases". Circulation Research. 116 (8): 1431–47. doi:10.1161/CIRCRESAHA.116.303311. PMC 4394208. PMID 25858067.
- 1 2 3 Perreau-Lenz S, Vengeliene V, Noori HR, Merlo-Pich EV, Corsi MA, Corti C, Spanagel R (August 2012). "Inhibition of the casein-kinase-1-ε/δ/ prevents relapse-like alcohol drinking". Neuropsychopharmacology. 37 (9): 2121–31. doi:10.1038/npp.2012.62. PMC 3398717. PMID 22549116.
- ↑ De Nobrega AK, Lyons LC (April 2016). "Circadian Modulation of Alcohol-Induced Sedation and Recovery in Male and Female Drosophila". Journal of Biological Rhythms. 31 (2): 142–60. doi:10.1177/0748730415627067. PMC 5136465. PMID 26833081.
- ↑ Goldberg LR, Kirkpatrick SL, Yazdani N, Luttik KP, Lacki OA, Babbs RK, Jenkins DF, Johnson WE, Bryant CD (September 2017). "Casein kinase 1-epsilon deletion increases mu opioid receptor-dependent behaviors and binge eating1". Genes, Brain and Behavior. 16 (7): 725–738. doi:10.1111/gbb.12397. PMC 6180211. PMID 28594147.
- ↑ Zhang Y, Qiu WJ, Chan SC, Han J, He X, Lin SC (May 2002). "Casein kinase I and casein kinase II differentially regulate axin function in Wnt and JNK pathways". The Journal of Biological Chemistry. 277 (20): 17706–12. doi:10.1074/jbc.M111982200. PMID 11884395.
- ↑ Smadja Storz S, Tovin A, Mracek P, Alon S, Foulkes NS, Gothilf Y (2013-01-21). "Casein kinase 1δ activity: a key element in the zebrafish circadian timing system". PLOS ONE. 8 (1): e54189. Bibcode:2013PLoSO...854189S. doi:10.1371/journal.pone.0054189. PMC 3549995. PMID 23349822.
- 1 2 Badura L, Swanson T, Adamowicz W, Adams J, Cianfrogna J, Fisher K, Holland J, Kleiman R, Nelson F, Reynolds L, St Germain K, Schaeffer E, Tate B, Sprouse J (August 2007). "An inhibitor of casein kinase I epsilon induces phase delays in circadian rhythms under free-running and entrained conditions". The Journal of Pharmacology and Experimental Therapeutics. 322 (2): 730–8. doi:10.1124/jpet.107.122846. PMID 17502429. S2CID 85875627.
- 1 2 Kon N, Sugiyama Y, Yoshitane H, Kameshita I, Fukada Y (2015-07-25). "Cell-based inhibitor screening identifies multiple protein kinases important for circadian clock oscillations". Communicative & Integrative Biology. 8 (4): e982405. doi:10.4161/19420889.2014.982405. PMC 4594307. PMID 26478783.
- ↑ Knippschild U, Krüger M, Richter J, Xu P, García-Reyes B, Peifer C, Halekotte J, Bakulev V, Bischof J (2014). "The CK1 Family: Contribution to Cellular Stress Response and Its Role in Carcinogenesis". Frontiers in Oncology. 4: 96. doi:10.3389/fonc.2014.00096. PMC 4032983. PMID 24904820.