| Gyroporus cyanescens | |
|---|---|
 | |
| Scientific classification | |
| Domain: | Eukaryota |
| Kingdom: | Fungi |
| Division: | Basidiomycota |
| Class: | Agaricomycetes |
| Order: | Boletales |
| Family: | Gyroporaceae |
| Genus: | Gyroporus |
| Species: | G. cyanescens |
| Binomial name | |
| Gyroporus cyanescens | |
| Synonyms[1] | |
|
Boletus cyanescens Bull. (1788) | |
| Gyroporus cyanescens | |
|---|---|
| Pores on hymenium | |
| Cap is convex or flat | |
| Hymenium attachment is irregular or not applicable | |
| Stipe is bare | |
| Spore print is cream to yellow | |
| Ecology is mycorrhizal | |
| Edibility is choice | |
Gyroporus cyanescens, commonly known as the bluing bolete or the cornflower bolete, is a species of bolete fungus in the family Gyroporaceae. First described from France in 1788, the species is found in Asia, Australia, Europe, and eastern North America, where it grows on the ground in coniferous and mixed forests.
The yellowish to buff cap surface is fibrous and roughened, and reaches up to 12 cm (4.7 in) in diameter. The thick stem, roughly the same color as the cap or lighter, is hollowed out into chambers. All parts of the mushroom turn an intense blue color within a few moments of bruising or cutting. The mushroom is edible, despite its hard stem. A less common variety occurs where the color change is to deep violet rather than blue. The bluing reaction results from the oxidation of a chemical called gyrocyanin.
Taxonomy
The species was first described scientifically by French botanist Jean Baptiste François Pierre Bulliard in his 1788 Herbier de la France.[2] Later synonyms include Boletus constrictus by Christian Hendrik Persoon in 1801,[3] Leccinum constrictum by Samuel Frederick Gray in 1821,[4] Suillus cyanescens by Petter Karsten in 1882,[5] and Leucoconius cyanescens by Günther Beck von Mannagetta und Lerchenau in 1923.[6] The variety violaceotinctus was described by Roy Watling in 1969 from collections made in Michigan, USA.[7]
The specific epithet is derived from the Ancient Greek κύανoς, meaning "dark blue",[8] while the varietal epithet violaceotinctus means "having a violet tinge".[9] It is commonly known as the bluing bolete[10] or the cornflower bolete.[11]
Description
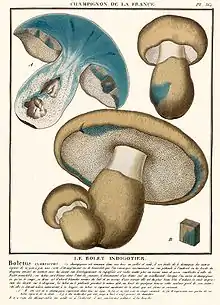
The cap of G. cyanescens is initially convex, but flattens out in maturity, sometimes becoming shallowly depressed;[11] it reaches a diameter of 4–12 cm (1.6–4.7 in). The cap is dry, and ranges in color from buff to yellowish to pale olive, occasionally with darker streaks of color.[10] Its surface is uneven, sometimes with wrinkles and pits.[11] The cap margin is initially curved inward, and sometimes splits in maturity.[12] The flesh is whitish to pale yellow, and has a brittle texture. On the underside of the cap, the pore surface is white to yellowish, sometimes with olive or tan tinges. There are roughly two circular pores per millimeter, and the tubes that comprise the pores are 5–10 mm (0.2–0.4 in) deep, but depressed around the top of the stem. Injury to the pores will cause them to stain first greenish yellow, then greenish blue or blue.[10]

The stem is 4–10 cm (1.6–3.9 in) long by 1–2.5 cm (0.4–1.0 in) thick, and is either roughly equal in width throughout its length, or has a basal or middle swelling. The stem tissue is hard and brittle; it is initially stuffed with a soft pith that develops cavities, or becomes entirely hollow in maturity. Roughly the same color as the cap or lighter, the stem surface is dry and lacks reticulations. It is initially covered in coarse hairs that tend to disappear in maturity to leave a relatively smooth surface. All parts of the fruit body stain blue when cut or injured. The variety G. cyanescens var. violaceotinctus is nearly identical in appearance, but stains dark lilac to indigo when bruised.[10] The odor and taste of the fruit bodies is indistinct.[13]
The color of the spore print is pale yellow. Spores are ellipsoid, smooth, hyaline (translucent), and have dimensions of 8–10 by 5–6 µm.[10] The basidia (spore-bearing cells) are club shaped, two- to four-spored, and measure 24–30 by 8–10 µm. Pleurocystidia (cystidia on the inner walls of the tubes) are light yellow brown in color, club shaped, infrequent, and measure 25–38 by 7.2 µm; the cheilocystidia (found on the tube edge) are colorless, numerous, and measure 32–47 by 7–10 µm. Clamp connections are present in the hyphae.[14]
Similar species
Although there are a few lookalike species with similar overall appearance, in the field, Gyroporus cyanescens is typically readily recognized by its characteristic straw-yellow color and nearly instantaneous dark blue bruising. G. phaeocyanescens is smaller, with a dull brownish-yellow cap. Although its flesh has a bluing reaction to injury, its yellow pore surface does not. It has larger spores, measuring 9–15 by 5–7 µm. G. umbrinosquamosus, found along the Gulf Coast of the United States, is similar in appearance, but lacks the bluing reaction.[10] Newly described from China in 2003, G. brunneofloccosus closely resembles G. cyanescens, and was frequently confused with that species. It has a smaller fruit body, with a brownish cap up to 8 cm (3.1 in) in diameter. Its staining reaction involves a change from light turquoise to dark turquoise or dark blue. Its spores are 5–8.5 by 4–5.3 µm.[15] Suillus tomentosus has brownish pores that undergo a slower blue staining reaction.[11] If the fruit bodies are not uprooted and only the top of the cap is examined, G. cyanescens can be confused with young Russula fellea mushrooms.[16]
Uses
Gyroporus cyanescens is edible, and considered "choice" by several sources.[17][18][19] The fruit bodies, even if mature, are typically free of insect larvae. Specimens collected in sandy soil, however, are difficult to clean,[9] but cleaning may be facilitated by washing the fruit bodies in a bowl of water so that the sand sinks to the bottom. The blue color largely disappears after two minutes of sautéing. Cooked mushrooms have a meaty texture, and mild nutty flavor that is enhanced if fried to crispness. Drying the mushrooms strengthens the taste.[19]
The variety violaceotinctus is used in mushroom dyeing, and produces a light yellow, beige, gold, or brownish-orange color depending on the mordant used.[20]
Ecology and distribution
Gyroporus cyanescens is an ectomycorrhizal species that has a broad host range.[21] Fruit bodies of Gyroporus cyanescens grow singly or scattered on the ground in deciduous and mixed forests. Often found in association with birch and poplar, the fungus tends to prefer sandy soil,[10] and also frequents road banks and woodland edges.[22] Fruiting occurs in summer and early autumn.[18] Fruit bodies can be parasitized by the mold Sepedonium ampullosporum.[23] Infection results in necrosis of the mushroom tissue, and a yellow color caused by the formation of large amounts of pigmented aleurioconidia (single-celled conidia produced by extrusion from the conidiophores).[24]
Gyroporus cyanescens is found in Asia, Australia, North America, and Europe. In China, it is known from Guangdong and Yunnan.[14] The fungus appears in eucalypt woodland in Australia.[25] In North America, it is widespread east of the Rocky Mountains.[13] The geographical distribution ranges from eastern Canada to Florida, and west to Minnesota,[10] although it has been occasionally reported from the Pacific Northwest.[19] G. cyanescens var. violaceotinctus has been reported from Japan.[26]
Chemistry

The identity of the chemical causing bluing upon tissue injury was reported in 1973.[27] The molecule, gyrocyanin, is a highly oxidized bis-phenol-substituted cyclopentenone that develops a blue color when it is oxidized. In contrast, the bluing of other boletes has been attributed to the oxidation of variegatic or xerocomic acid.[28] Gyrocyanin is biosynthesized from intermediates supplied by the shikimate pathway, a metabolic route used by fungi for the synthesis of aromatic amino acids.[29]
See also
References
- ↑ "Gyroporus cyanescens (Bull.) Quél. 1886". MycoBank. International Mycological Association. Retrieved 2012-07-22.
- ↑ Bulliard JBF. (1788). Herbier de la France (in French). Vol. 8. Paris, France: Chez l'auteur, Didot, Debure, Belin. p. plate 369. doi:10.5962/bhl.title.5365.
- ↑ Persoon CH. (1801). Synopsis Methodica Fungorum (in Latin). Göttingen, Sweden: H. Dieterich. p. 508.
- ↑ Gray SF. (1821). A Natural Arrangement of British Plants. Vol. 1. London, UK: Baldwin, Cradock, and Joy. p. 647. doi:10.5962/bhl.title.43804.
- ↑ Karsten P. (1882). "Rysslands, Finlans och den Skandinaviska halföns Hattsvampar. Sednare Delen: Pip-, Tagg-, Hud-, Klubb- och Gelésvampar". Bidrag till Kännedom of Finlands Natur Folk (in Swedish). 37: 1.
- ↑ Beck G. (1923). "Versuch einer systematischen Gliederung der Gattung Boletus L. em". Zeitschrift für Pilzkunde (in German). 2 (7): 141–9.
- ↑ Watling R. (1969). "New fungi from Michigan USA. Paxillus vernalis new species Gyroporus cyanescens var violaceotinctus new variety". Notes from the Royal Botanic Garden Edinburgh. 29 (1): 59–66. ISSN 0080-4274.
- ↑ Rea C. (1922). British Basidiomycetae: A Handbook to the Larger British Fungi. Cambridge, UK: Cambridge University Press. p. 553.
- 1 2 Smith AH, Weber NS (1980). The Mushroom Hunter's Field Guide. Ann Arbor, Michigan: University of Michigan Press. p. 59. ISBN 0-472-85610-3.
- 1 2 3 4 5 6 7 8 Bessette AE, Roody WC, Bessette AR (2000). North American Boletes. Syracuse, New York: Syracuse University Press. pp. 189–91. ISBN 9780815605881.
- 1 2 3 4 McKnight VB, McKnight KH (1987). A Field Guide to Mushrooms: North America. Peterson Field Guides. Boston, Massachusetts: Houghton Mifflin. p. 110. ISBN 0-395-91090-0.
- ↑ Phillips R. (2005). Mushrooms and Other Fungi of North America. Buffalo, New York: Firefly Books. p. 274. ISBN 1-55407-115-1.
- 1 2 Kuo M. (March 2003). "Gyroporus cyanescens". MushroomExpert.com. Retrieved 2012-07-22.
- 1 2 Zhishu B, Zheng G, Taihui L (1993). The Macrofungus Flora of China's Guangdong Province (Chinese University Press). New York, New York: Columbia University Press. p. 468. ISBN 962-201-556-5.
- ↑ Li TH, Deng WQ, Song B (2003). "A new cyanescent species of Gyroporus from China" (PDF). Fungal Diversity. 12: 123–7.
- ↑ Mattock G. (1999). "Gyroporus cyanescens: easily overlooked in Hampshire". Mycologist. 13 (2): 64. doi:10.1016/S0269-915X(99)80009-3.
- ↑ Bessette A, Fischer DH (1992). Edible Wild Mushrooms of North America: A Field-to-Kitchen Guide. Austin, Texas: University of Texas Press. pp. 100–2. ISBN 0-292-72080-7.
- 1 2 Miller HR, Miller OK (2006). North American Mushrooms: A Field Guide to Edible and Inedible Fungi. Guilford, Connecticut: Falcon Guide. p. 367. ISBN 0-7627-3109-5.
- 1 2 3 Kuo M. (2007). 100 Edible Mushrooms. Ann Arbor, Michigan: The University of Michigan Press. pp. 169–71. ISBN 978-0-472-03126-9.
- ↑ Bessette A, Bessette AR (2001). The Rainbow Beneath my Feet: A Mushroom Dyer's Field Guide. Syracuse, New York: Syracuse University Press. p. 47. ISBN 0-8156-0680-X.
- ↑ Smith SE, Read DJ (2008). Mycorrhizal Symbiosis. Academic Press. p. 198. ISBN 978-0-12-370526-6.
- ↑ Roody WC. (2003). Mushrooms of West Virginia and the Central Appalachians. Lexington, Kentucky: University Press of Kentucky. p. 322. ISBN 0-8131-9039-8.
- ↑ Ammer H, Besl H, Vilsmeier S (1997). "Sepedonium ampullosporum, a thermophilic hyphomycete parasitizing on fruit-bodies of Boletales". Zeitschrift für Mykologie (in German). 5 (2): 127–32. ISSN 0170-110X.
- ↑ Neuhof T, Berg A, Besl H, Schweche T, Dieckmann R, von Döhren H (2007). "Peptaibol production by Sepedonium strains parasitizing Boletales". Chemistry & Biodiversity. 4 (6): 1103–15. doi:10.1002/cbdv.200790099. PMID 17589879. S2CID 40865857.
- ↑ Hilton RN. (1972). "Mycology in Western Australia". Bulletin of the British Mycological Society. 7 (1): 27–8. doi:10.1016/S0007-1528(73)80030-6.
- ↑ Nagasawa E. (2001). "Taxonomic studies of Japanese boletes. I. The genera Boletinellus, Gyrodon and Gyroporus". Reports of the Tottori Mycological Institute (in Japanese). 1 (39): 1–27. ISSN 0388-8266.
- ↑ Besl H, Bresinsky A, Steglish W, Zipfel K (1973). "Pilzpigmente, XVII. Über Gyrocyanin, das blauende Prinzip des Kornblumenröhrlings (Gyroporus cyanescens), und eine oxidative Ringverengung des Atromentins" [Fungus pigments .17. Gyrocyanin, blueing principle of Gyroporus cyanescens, and an oxidative ring contraction of atromentin]. Chemische Berichte/Recueil (in German). 106 (10): 3223–9. doi:10.1002/cber.19731061012.
- ↑ Nelson SF. (2010). "Bluing components and other pigments of Boletes" (PDF). FUNGI. 3 (4): 11–4.
- ↑ Steglich W. (1975). "Pilzarbstoffe". Chemie in unserer Zeit (in German). 9 (4): 117–23. doi:10.1002/ciuz.19750090404.
External links
- Gyroporus cyanescens in Index Fungorum
- Tom Volk's Fungus of the Month for July 2003
- Youtube Video of staining reaction
- Image of spores