Induced stem cells (iSC) are stem cells derived from somatic, reproductive, pluripotent or other cell types by deliberate epigenetic reprogramming. They are classified as either totipotent (iTC), pluripotent (iPSC) or progenitor (multipotent – iMSC, also called an induced multipotent progenitor cell – iMPC) or unipotent – (iUSC) according to their developmental potential and degree of dedifferentiation. Progenitors are obtained by so-called direct reprogramming or directed differentiation and are also called induced somatic stem cells.
Three techniques are widely recognized:[1]
- Transplantation of nuclei taken from somatic cells into an oocyte (egg cell) lacking its own nucleus (removed in lab)[2][3][4][5]
- Fusion of somatic cells with pluripotent stem cells[6] and
- Transformation of somatic cells into stem cells, using the genetic material encoding reprogramming protein factors,[7][8][9] recombinant proteins;[10] microRNA,[11][12][13][14][15] a synthetic, self-replicating polycistronic RNA[16] and low-molecular weight biologically active substances.[17][18][19]
Natural processes
In 1895 Thomas Morgan removed one of a frog's two blastomeres and found that amphibians are able to form whole embryos from the remaining part. This meant that the cells can change their differentiation pathway. In 1924 Hans Spemann and Hilde Mangold demonstrated the key importance of cell–cell inductions during animal development.[20] The reversible transformation of cells of one differentiated cell type to another is called metaplasia.[21] This transition can be a part of the normal maturation process, or caused by an inducement.
One example is the transformation of iris cells to lens cells in the process of maturation and transformation of retinal pigment epithelium cells into the neural retina during regeneration in adult newt eyes. This process allows the body to replace cells not suitable to new conditions with more suitable new cells. In Drosophila imaginal discs, cells have to choose from a limited number of standard discrete differentiation states. The fact that transdetermination (change of the path of differentiation) often occurs for a group of cells rather than single cells shows that it is induced rather than part of maturation.[22]
The researchers were able to identify the minimal conditions and factors that would be sufficient for starting the cascade of molecular and cellular processes to instruct pluripotent cells to organize the embryo. They showed that opposing gradients of bone morphogenetic protein (BMP) and Nodal, two transforming growth factor family members that act as morphogens, are sufficient to induce molecular and cellular mechanisms required to organize, in vivo or in vitro, uncommitted cells of the zebrafish blastula animal pole into a well-developed embryo.[23]
Some types of mature, specialized adult cells can naturally revert to stem cells. For example, "chief" cells express the stem cell marker Troy. While they normally produce digestive fluids for the stomach, they can revert into stem cells to make temporary repairs to stomach injuries, such as a cut or damage from infection. Moreover, they can make this transition even in the absence of noticeable injuries and are capable of replenishing entire gastric units, in essence serving as quiescent "reserve" stem cells.[24] Differentiated airway epithelial cells can revert into stable and functional stem cells in vivo.[25] After injury, mature terminally differentiated kidney cells dedifferentiate into more primordial versions of themselves and then differentiate into the cell types needing replacement in the damaged tissue[26] Macrophages can self-renew by local proliferation of mature differentiated cells.[27][28] In newts, muscle tissue is regenerated from specialized muscle cells that dedifferentiate and forget the type of cell they had been. This capacity to regenerate does not decline with age and may be linked to their ability to make new stem cells from muscle cells on demand.[29]
A variety of nontumorigenic stem cells display the ability to generate multiple cell types. For instance, multilineage-differentiating stress-enduring (Muse) cells are stress-tolerant adult human stem cells that can self-renew. They form characteristic cell clusters in suspension culture that express a set of genes associated with pluripotency and can differentiate into endodermal, ectodermal and mesodermal cells both in vitro and in vivo.[30][31][32][33][34]
Other well-documented examples of transdifferentiation and their significance in development and regeneration were described in detail.[35][36]

Induced totipotent cells
SCNT-mediated
Induced totipotent cells can be obtained by reprogramming somatic cells with somatic-cell nuclear transfer (SCNT). The process involves sucking out the nucleus of a somatic (body) cell and injecting it into an oocyte that has had its nucleus removed[3][5][37][38][39][40]
Using an approach based on the protocol outlined by Tachibana et al.,[3] hESCs can be generated by SCNT using dermal fibroblasts nuclei from both a middle-aged 35-year-old male and an elderly, 75-year-old male, suggesting that age-associated changes are not necessarily an impediment to SCNT-based nuclear reprogramming of human cells.[41] Such reprogramming of somatic cells to a pluripotent state holds huge potentials for regenerative medicine. Unfortunately, the cells generated by this technology are potentially not completely protected from the immune system of the patient (donor of nuclei), because they have the same mitochondrial DNA, as a donor of oocytes, instead of the patients mitochondrial DNA. This reduces their value as a source for autologous stem cell transplantation therapy; as for the present,[42] it is not clear whether it can induce an immune response of the patient upon treatment.
Induced androgenetic haploid embryonic stem cells can be used instead of sperm for cloning. These cells, synchronized in M phase and injected into the oocyte, can produce viable offspring.[43]
These developments, together with data on the possibility of unlimited oocytes from mitotically active reproductive stem cells,[44] offer the possibility of industrial production of transgenic farm animals. Repeated recloning of viable mice through a SCNT method that includes a histone deacetylase inhibitor, trichostatin, added to the cell culture medium,[45] show that it may be possible to reclone animals indefinitely with no visible accumulation of reprogramming or genomic errors[46] However, research into technologies to develop sperm and egg cells from stem cells raises bioethical issues.[47]
Such technologies may also have far-reaching clinical applications for overcoming cytoplasmic defects in human oocytes.[3][48] For example, the technology could prevent inherited mitochondrial disease from passing to future generations. Mitochondrial genetic material is passed from mother to child. Mutations can cause diabetes, deafness, eye disorders, gastrointestinal disorders, heart disease, dementia, and other neurological diseases. The nucleus from one human egg has been transferred to another, including its mitochondria, creating a cell that could be regarded as having two mothers. The eggs were then fertilised and the resulting embryonic stem cells carried the swapped mitochondrial DNA.[49] As evidence that the technique is safe, the author of this method points to the existence of the healthy monkeys that are now more than four years old – and are the product of mitochondrial transplants across different genetic backgrounds.[50]
In late-generation telomerase-deficient (Terc−/−) mice, SCNT-mediated reprogramming mitigates telomere dysfunction and mitochondrial defects to a greater extent than iPSC-based reprogramming.[51]
Other cloning and totipotent transformation achievements have been described.[52]
Obtained without SCNT
Recently, some researchers succeeded in getting totipotent cells without the aid of SCNT. Totipotent cells were obtained using epigenetic factors such as oocyte germinal isoform of histone.[53] Reprogramming in vivo, by transitory induction of the four factors Oct4, Sox2, Klf4 and c-Myc in mice, confers totipotency features. Intraperitoneal injection of such in vivo iPS cells generates embryo-like structures that express embryonic and extraembryonic (trophectodermal) markers.[54] The developmental potential of mouse pluripotent stem cells to yield both embryonic and extra-embryonic lineages also can be expanded by microRNA miR-34a deficiency, leading to strong induction of endogenous retroviruses MuERV-L (MERVL).[55]
Rejuvenation to iPSCs
Transplantation of pluripotent/embryonic stem cells into the body of adult mammals usually leads to the formation of teratomas, which can then turn into a malignant tumor teratocarcinoma. However, putting embryonic stem cells ot teratocarcinoma cells into the embryo at the blastocyst stage caused them to become incorporated in the cell mass and often produced a normal healthy chimeric (i.e. composed of cells from different organisms) animal.[56] iPSc were first obtained in the form of transplantable teratocarcinoma induced by grafts taken from mouse embryos.[57] Teratocarcinoma formed from somatic cells.[58] Genetically mosaic mice were obtained from malignant teratocarcinoma cells, confirming the cells' pluripotency.[59][60][61] It turned out that teratocarcinoma cells are able to maintain a culture of pluripotent embryonic stem cell in an undifferentiated state, by supplying the culture medium with various factors.[62] In the 1980s, it became clear that transplanting pluripotent/embryonic stem cells into the body of adult mammals, usually leads to the formation of teratomas, which can then turn into a malignant tumor teratocarcinoma.[63] However, putting teratocarcinoma cells into the embryo at the blastocyst stage caused them to become incorporated in the inner cell mass and often produced a normal chimeric (i.e. composed of cells from different organisms) animal.[64][65][66] This indicated that the cause of the teratoma is a dissonance - mutual miscommunication between young donor cells and surrounding adult cells (the recipient's so-called "niche").
In August 2006, Japanese researchers circumvented the need for an oocyte, as in SCNT. By reprograming mouse embryonic fibroblasts into pluripotent stem cells via the ectopic expression of four transcription factors, namely Oct4, Sox2, Klf4 and c-Myc, they proved that the overexpression of a small number of factors can push the cell to transition to a new stable state that is associated with changes in the activity of thousands of genes.[7]

Reprogramming mechanisms are thus linked, rather than independent, and are centered on a small number of genes.[67] IPSC properties are very similar to ESCs.[68] iPSCs have been shown to support the development of all-iPSC mice using a tetraploid (4n) embryo,[69] the most stringent assay for developmental potential. However, some genetically normal iPSCs failed to produce all-iPSC mice because of aberrant epigenetic silencing of the imprinted Dlk1-Dio3 gene cluster.[18] A team headed by Hans Schöler (who discovered the Oct4 gene back in 1989) showed that Oct4 overexpression drives massive off-target gene activation during reprogramming deteriorating the quality of iPSCs. Compared to OSKM (Oct4, Sox2, Klf4 and c-Myc) that show abnormal imprinting and differentiation patterns, SKM (Sox2, Klf4 and c-Myc) reprogramming generates iPSCs with high developmental potential (nearly 20-fold higher than that of OSKM) equivalent to embryonic stem cell, as determined by their ability to generate all-iPSC mice through tetraploid embryo complementation.[70]
An important advantage of iPSC over ESC is that they can be derived from adult cells, rather than from embryos. Therefore, it becomes possible to obtain iPSC from adult and even elderly patients.[9][71][72]
Reprogramming somatic cells into iPSCs leads to rejuvenation. It was found that reprogramming leads to telomere lengthening and subsequent shortening after their differentiation back into fibroblast-like derivatives.[73] Thus, reprogramming leads to the restoration of embryonic telomere length,[74] and hence increases the potential number of cell divisions otherwise limited by the Hayflick limit.[75]
However, because of the dissonance between rejuvenated cells and the surrounding niche of the recipient's older cells, the injection of his own iPSC usually leads to an immune response,[76] which can be used for medical purposes,[77] or the formation of tumors such as teratoma.[78] A hypothesized reason is that some cells differentiated from ESCs and iPSCs in vivo continue to synthesize embryonic protein isoforms.[79] So, the immune system might detect and attack cells that are not cooperating properly.
A small molecule called MitoBloCK-6 can force the pluripotent stem cells to die by triggering apoptosis (via cytochrome c release across the mitochondrial outer membrane) in human pluripotent stem cells, but not in differentiated cells. Shortly after differentiation, daughter cells became resistant to death. When MitoBloCK-6 was introduced to differentiated cell lines, the cells remained healthy. The key to their survival was hypothesized to be due to the changes undergone by pluripotent stem cell mitochondria in the process of cell differentiation. This ability of MitoBloCK-6 to separate the pluripotent and differentiated cell lines has the potential to reduce the risk of teratomas and other problems in regenerative medicine.[80]
In 2012, other small molecules (selective cytotoxic inhibitors of human pluripotent stem cells – hPSCs) were identified that prevented human pluripotent stem cells from forming teratomas in mice. The most potent and selective compound of them (PluriSIn #1) inhibits stearoyl-coA desaturase (the key enzyme in oleic acid biosynthesis), which finally results in apoptosis. With the help of this molecule, the undifferentiated cells can be selectively removed from culture.[81][82] An efficient strategy to selectively eliminate pluripotent cells with teratoma potential is targeting pluripotent stem cell-specific antiapoptotic factor(s) (i.e., survivin or Bcl10). A single treatment with chemical survivin inhibitors (e.g., quercetin or YM155) can induce selective and complete cell death of undifferentiated hPSCs and is claimed to be sufficient to prevent teratoma formation after transplantation.[83] However, it is unlikely that any kind of preliminary clearance is able to secure the replanting of iPSCs or ESCs. After the selective removal of pluripotent cells, they re-emerge quickly by reverting differentiated cells into stem cells, which leads to tumors.[84] This may be due to the disorder of let-7 regulation of its target Nr6a1 (also known as Germ cell nuclear factor - GCNF), an embryonic transcriptional repressor of pluripotency genes that regulates gene expression in adult fibroblasts following micro-RNA miRNA loss.[85]
Teratoma formation by pluripotent stem cells may be caused by low activity of PTEN enzyme, reported to promote the survival of a small population (0.1–5% of total population) of highly tumorigenic, aggressive, teratoma-initiating embryonic-like carcinoma cells during differentiation. The survival of these teratoma-initiating cells is associated with failed repression of Nanog, as well as a propensity for increased glucose and cholesterol metabolism.[86] These teratoma-initiating cells also expressed a lower ratio of p53/p21 when compared to non-tumorigenic cells.[87] In connection with the above safety problems, the use of iPSCs for cell therapy is still limited.[88] However, they can be used for a variety of other purposes, including the modeling of disease,[89] screening (selective selection) of drugs, and toxicity testing of various drugs.[90]

The tissue grown from iPSCs, placed in the "chimeric" embryos in the early stages of mouse development, practically do not cause an immune response (after the embryos have grown into adult mice) and are suitable for autologous transplantation[91] At the same time, full reprogramming of adult cells in vivo within tissues by transitory induction of the four factors Oct4, Sox2, Klf4 and c-Myc in mice results in teratomas emerging from multiple organs.[54] Furthermore, partial reprogramming of cells toward pluripotency in vivo in mice demonstrates that incomplete reprogramming entails epigenetic changes (failed repression of Polycomb targets and altered DNA methylation) in cells that drive cancer development.[92] However, a number of researchers later managed to carry out cyclical partial reprogramming in vivo by expression of the Yamanaka factors for a short period of time without subsequent carcinogenesis, thus partially rejuvenating and extending lifespan in physiologically aging wild-type mice[93] and progeroid mice.[94] With in vitro method employing slightly longer periods of reprogramming (to substantially rejuvenate) cells temporarily lose their cell identity[95] but "reacquire their initial somatic fate when the reprogramming factors are withdrawn".[96][97][98]
Chemical inducement
By using solely small molecules, Deng Hongkui and colleagues demonstrated that endogenous "master genes" are enough for cell fate reprogramming. They induced a pluripotent state in adult cells from mice using seven small-molecule compounds.[17] The method's effectiveness is quite high: it was able to convert 0.02% of the adult tissue cells into iPSCs, which is comparable to the gene insertion conversion rate. The authors note that the mice generated from CiPSCs were "100% viable and apparently healthy for up to 6 months". So, this chemical reprogramming strategy has potential use in generating functional desirable cell types for clinical applications.[100][101]
In 2015 a robust chemical reprogramming system was established with a yield up to 1,000-fold greater than that of the previously reported protocol. So, chemical reprogramming became a promising approach to manipulate cell fates.[102]
Differentiation from induced teratoma
The fact that human iPSCs are capable of forming teratomas not only in humans but also in some animal body, in particular mice or pigs, allowed researchers to develop a method for differentiation of iPSCs in vivo. For this purpose, iPSCs with an agent for inducing differentiation into target cells are injected to a genetically modified pig or mouse that has suppressed immune system activation on human cells. The formed teratoma is cut out and used for the isolation of the necessary differentiated human cells[103] by means of monoclonal antibody to tissue-specific markers on the surface of these cells. This method has been successfully used for the production of functional myeloid, erythroid, and lymphoid human cells suitable for transplantation (yet only to mice).[104] Mice engrafted with human iPSC teratoma-derived hematopoietic cells produced human B and T cells capable of functional immune responses. These results offer hope that in vivo generation of patient customized cells is feasible, providing materials that could be useful for transplantation, human antibody generation, and drug screening applications. Using MitoBloCK-6[80] and/or PluriSIn # 1 the differentiated progenitor cells can be further purified from teratoma forming pluripotent cells. The fact that the differentiation takes place even in the teratoma niche offers hope that the resulting cells are sufficiently stable to stimuli able to cause their transition back to the dedifferentiated (pluripotent) state and therefore safe. A similar in vivo differentiation system, yielding engraftable hematopoietic stem cells from mouse and human iPSCs in teratoma-bearing animals in combination with a maneuver to facilitate hematopoiesis, was described by Suzuki et al.[105] They noted that neither leukemia nor tumors were observed in recipients after intravenous injection of iPSC-derived hematopoietic stem cells into irradiated recipients. Moreover, this injection resulted in multilineage and long-term reconstitution of the hematolymphopoietic system in serial transfers. Such a system provides a useful tool for practical application of iPSCs in the treatment of hematologic and immunologic diseases.[106]
For further development of this method, animals (such as a mice) in which the human cell graft is grown must have modified genome such that all its cells express and have on its surface human SIRPα.[107] To prevent rejection after transplantation to the patient of the allogenic organ or tissue, grown from the pluripotent stem cells in vivo in the animal, these cells should express two molecules: CTLA4-Ig, which disrupts T cell costimulatory pathways and PD-L1, which activates T cell inhibitory pathway.[108]
See also: US 20130058900 patent.
Differentiated cell types
Retinal cells
In the near-future, clinical trials designed to demonstrate the safety of the use of iPSCs for cell therapy of people with age-related macular degeneration, a disease causing blindness through retina damaging, will begin. There are several articles describing methods for producing retinal cells from iPSCs[109][110] and how to use them for cell therapy.[111][112] Reports of iPSC-derived retinal pigmented epithelium transplantation showed enhanced visual-guided behaviors of experimental animals for 6 weeks after transplantation.[113] However, clinical trials have been successful: ten patients with retinitis pigmentosa have had eyesight restored, including a woman who had only 17 percent of her vision left.[114]
Lung and airway epithelial cells
Chronic lung diseases, such as idiopathic pulmonary fibrosis and cystic fibrosis or chronic obstructive pulmonary disease and asthma, are leading causes of morbidity and mortality worldwide, with a considerable human, societal, and financial burden. Therefore, there is an urgent need for effective cell therapy and lung tissue engineering.[115][116] Several protocols have been developed for generation of most cell types of the respiratory system, which may be useful for deriving patient-specific therapeutic cells.[117][118][119][120][121]
Reproductive cells
Some lines of iPSCs have the potential to differentiate into male germ cells and oocyte-like cells in an appropriate niche (by culturing in retinoic acid and porcine follicular fluid differentiation medium or seminiferous tubule transplantation). Moreover, iPSC transplantation contributes to repairing the testis of infertile mice, demonstrating the potential of gamete derivation from iPSCs in vivo and in vitro.[122]
Induced progenitor stem cells
Direct transdifferentiation
The risk of cancer and tumors creates the need to develop methods for safer cell lines suitable for clinical use. An alternative approach is so-called "direct reprogramming" – transdifferentiation of cells without passing through the pluripotent state.[123][124][125][126][127][128][129] The basis for this approach was that 5-azacytidine – a DNA demethylation reagent – can cause the formation of myogenic, chondrogenic, and adipogeni clones in an immortal cell line of mouse embryonic fibroblasts[130] and that the activation of a single gene, later named MyoD1, is sufficient for such reprogramming.[131] Compared with iPSCs whose reprogramming requires at least two weeks, the formation of induced progenitor cells sometimes occurs within a few days and the efficiency of reprogramming is usually many times higher. This reprogramming does not always require cell division.[132] The cells resulting from such reprogramming are more suitable for cell therapy because they do not form teratomas.[128] For example, Chandrakanthan et al., & Pimanda describe the generation of tissue-regenerative multipotent stem cells (iMS cells) by treating mature bone and fat cells transiently with a growth factor (platelet-derived growth factor–AB (PDGF-AB)) and 5-Azacytidine. These authors state that "Unlike primary mesenchymal stem cells, which are used with little objective evidence in clinical practice to promote tissue repair, iMS cells contribute directly to in vivo tissue regeneration in a context-dependent manner without forming tumors" and thus "has significant scope for application in tissue regeneration".[133][134][135]
Single transcription factor transdifferentiation
Originally, only early embryonic cells could be coaxed into changing their identity. Mature cells are resistant to changing their identity once they've committed to a specific kind. However, brief expression of a single transcription factor, the ELT-7 GATA factor, can convert the identity of fully differentiated, specialized non-endodermal cells of the pharynx into fully differentiated intestinal cells in intact larvae and adult roundworm Caenorhabditis elegans with no requirement for a dedifferentiated intermediate.[136]
Transdifferentiation with CRISPR-mediated activator
The cell fate can be effectively manipulated by epigenome editing, in particular via the direct activation of specific endogenous gene expression with CRISPR-mediated activator. When dCas9 (which has been modified so that it no longer cuts DNA, but still can be guided to specific sequences and to bind to them) is combined with transcription activators, it can precisely manipulate endogenous gene expression. Using this method, Wei et al., enhanced the expression of endogenous Cdx2 and Gata6 genes by CRISPR-mediated activators, thus directly converting mouse embryonic stem cells into two extraembryonic lineages, i.e., typical trophoblast stem cells and extraembryonic endoderm cells.[137] An analogous approach was used to induce activation of the endogenous Brn2, Ascl1, and Myt1l genes to convert mouse embryonic fibroblasts to induced neuronal cells.[138] Thus, transcriptional activation and epigenetic remodeling of endogenous master transcription factors are sufficient for conversion between cell types. The rapid and sustained activation of endogenous genes in their native chromatin context by this approach may facilitate reprogramming with transient methods that avoid genomic integration and provides a new strategy for overcoming epigenetic barriers to cell fate specification.
Phased process modeling regeneration
Another way of reprogramming is the simulation of processes that occur during amphibian limb regeneration. In urodele amphibians, an early step in limb regeneration is skeletal muscle fiber dedifferentiation into a cellulate that proliferates into limb tissue. However, sequential small molecule treatment of the muscle fiber with myoseverin, reversine (the aurora B kinase inhibitor), and some other chemicals (BIO (glycogen synthase-3 kinase inhibitor), lysophosphatidic acid (pleiotropic activator of G-protein-coupled receptors), SB203580 (p38 MAP kinase inhibitor), or SQ22536 (adenylyl cyclase inhibitor)) causes the formation of new muscle cell types as well as other cell types such as precursors to fat, bone, and nervous system cells.[139]
Antibody-based transdifferentiation
The researchers discovered that GCSF-mimicking antibody can activate a growth-stimulating receptor on marrow cells in a way that induces marrow stem cells which normally develop into white blood cells to become neural progenitor cells. The technique[140] enables researchers to search large libraries of antibodies and quickly select the ones with a desired biological effect.[141][142]
Reprograming by bacteria
The human gastrointestinal tract is colonized by a vast community of symbionts and commensals. The researchers demonstrate the phenomenon of somatic cell reprograming by bacteria and generation of multipotential cells from adult human dermal fibroblast cells by incorporating Lactic acid bacteria [143] This cellular transdifferentiation is caused by ribosomes and "can occur via donor bacteria that are swallowed and digested by host cells, which may induce ribosomal stress and stimulate cellular developmental plasticity".[144]
Conditionally reprogrammed cells (CRC)
Schlegel and Liu[145] demonstrated that the combination of feeder cells[146][147][148] and a Rho kinase inhibitor (Y-27632) [149][150] induces normal and tumor epithelial cells from many tissues to proliferate indefinitely in vitro.[151] This process occurs without the need for transduction of exogenous viral or cellular genes. These cells have been termed "conditionally reprogrammed cells" (CRC).[152] The induction of CRCs is rapid and results from reprogramming of the entire cell population. CRCs do not express high levels of proteins characteristic of iPSCs or embryonic stem cells (ESCs) (e.g., Sox2, Oct4, Nanog, or Klf4). This induction of CRCs is reversible and removal of Y-27632 and feeders allows the cells to differentiate normally.[145][153][154] CRC technology can generate 2×106 cells in 5 to 6 days from needle biopsies and can generate cultures from cryopreserved tissue and from fewer than four viable cells. CRCs retain a normal karyotype and remain nontumorigenic. This technique also efficiently establishes cell cultures from human and rodent tumors.[145][155][156]
The ability to rapidly generate many tumor cells from small biopsy specimens and frozen tissue provides significant opportunities for cell-based diagnostics and therapeutics (including chemosensitivity testing) and greatly expands the value of biobanking.[157][145][155][156] Using CRC technology, researchers were able to identify an effective therapy for a patient with a rare type of lung tumor.[158] Engleman's group[159] describes a pharmacogenomics platform that facilitates fast discovery of drug combinations which can overcome resistance using CRC system. In addition, the CRC method allows for the genetic manipulation of epithelial cells ex vivo and their subsequent evaluation in vivo in the same host. While initial studies revealed that co-culturing epithelial cells with Swiss 3T3 cells J2 was essential for CRC induction, with transwell culture plates, physical contact between feeders and epithelial cells isn't required for inducing CRCs, and more importantly, irradiation of the feeder cells is required for this induction. Consistent with the transwell experiments, conditioned medium induces and maintains CRCs, which is accompanied by a concomitant increase of cellular telomerase activity. The activity of the conditioned medium correlates directly with radiation-induced feeder cell apoptosis. Thus, conditional reprogramming of epithelial cells is mediated by a combination of Y-27632 and a soluble factor(s) released by apoptotic feeder cells.[160]
Riegel et al.[161] demonstrate that mouse ME cells, isolated from normal mammary glands or from mouse mammary tumor virus (MMTV)-Neu–induced mammary tumors, can be cultured indefinitely as conditionally reprogrammed cells (CRCs). Cell surface progenitor-associated markers are rapidly induced in normal mouse ME-CRCs relative to ME cells. However, the expression of certain mammary progenitor subpopulations, such as CD49f+ ESA+ CD44+, drops significantly in later passages. Nevertheless, mouse ME-CRCs grown in a three-dimensional extracellular matrix gave rise to mammary acinar structures. ME-CRCs isolated from MMTV-Neu transgenic mouse mammary tumors express high levels of HER2/neu, as well as tumor-initiating cell markers, such as CD44+, CD49f+, and ESA+ (EpCam). These patterns of expression are sustained in later CRC passages. Early and late passage ME-CRCs from MMTV-Neu tumors that were implanted in the mammary fat pads of syngeneic or nude mice developed vascular tumors that metastasized within 6 weeks of transplantation. Importantly, the histopathology of these tumors was indistinguishable from that of the parental tumors that develop in the MMTV-Neu mice. Application of the CRC system to mouse mammary epithelial cells provides an attractive model system to study genetics and phenotype of normal and transformed mouse epithelium in a defined culture environment and for in vivo transplant studies.
A different approach to CRC is to inhibit CD47 – a membrane protein that is the thrombospondin-1 receptor. Loss of CD47 permits sustained proliferation of primary murine endothelial cells, increases asymmetric division, and enables these cells to spontaneously reprogram to form multipotent embryoid body-like clusters. CD47 knockdown acutely increases mRNA levels of c-Myc and other stem cell transcription factors in cells in vitro and in vivo. Thrombospondin-1 is a key environmental signal that inhibits stem cell self-renewal via CD47. Thus, CD47 antagonists enable cell self-renewal and reprogramming by overcoming negative regulation of c-Myc and other stem cell transcription factors.[162] In vivo blockade of CD47 using an antisense morpholino increases survival of mice exposed to lethal total body irradiation due to increased proliferative capacity of bone marrow-derived cells and radioprotection of radiosensitive gastrointestinal tissues.[163]
Lineage-specific enhancers
Differentiated macrophages can self-renew in tissues and expand long-term in culture.[27] Under certain conditions, macrophages can divide without losing features they have acquired while specializing into immune cells – which is usually not possible with differentiated cells. The macrophages achieve this by activating a gene network similar to one found in embryonic stem cells. Single-cell analysis revealed that, in vivo, proliferating macrophages can derepress a macrophage-specific enhancer repertoire associated with a gene network controlling self-renewal. This happened when concentrations of two transcription factors named MafB and c-Maf were naturally low or were inhibited for a short time. Genetic manipulations that turned off MafB and c-Maf in the macrophages caused the cells to start a self-renewal program. The similar network also controls embryonic stem cell self-renewal but is associated with distinct embryonic stem cell-specific enhancers.[28]
Hence macrophages isolated from MafB- and c-Maf-double deficient mice divide indefinitely; the self-renewal depends on c-Myc and Klf4.[164]
Indirect lineage conversion
Indirect lineage conversion is a reprogramming methodology in which somatic cells transition through a plastic intermediate state of partially reprogrammed cells (pre-iPSC), induced by brief exposure to reprogramming factors, followed by differentiation in a specially developed chemical environment (artificial niche).[165]
This method could be both more efficient and safer, since it does not seem to produce tumors or other undesirable genetic changes and results in much greater yield than other methods. However, the safety of these cells remains questionable. Since lineage conversion from pre-iPSC relies on the use of iPSC reprogramming conditions, a fraction of the cells could acquire pluripotent properties if they do not stop the de-differentation process in vitro or due to further de-differentiation in vivo.[166]
Outer membrane glycoprotein
A common feature of pluripotent stem cells is the specific nature of protein glycosylation of their outer membrane. That distinguishes them from most nonpluripotent cells, although not from white blood cells.[167] The glycans on the stem cell surface respond rapidly to alterations in cellular state and signaling and are therefore ideal for identifying even minor changes in cell populations. Many stem cell markers are based on cell surface glycan epitopes, including the widely used markers SSEA-3, SSEA-4, Tra 1-60 and Tra 1-81.[168] Suila Heli et al.[169] speculate that in human stem cells, extracellular O-GlcNAc and extracellular O-LacNAc play a crucial role in the fine tuning of Notch signaling pathway - a highly conserved cell signaling system that regulates cell fate specification, differentiation, left–right asymmetry, apoptosis, somitogenesis, and angiogenesis, and plays a key role in stem cell proliferation (reviewed by Perdigoto and Bardin[170] and Jafar-Nejad et al.[171])
Changes in outer membrane protein glycosylation are markers of cell states connected in some way with pluripotency and differentiation.[172] The glycosylation change is apparently not just the result of the initialization of gene expression, but also performs as an important gene regulator involved in the acquisition and maintenance of the undifferentiated state.[173]
For example, activation of glycoprotein ACA,[174] linking glycosylphosphatidylinositol on the surface of the progenitor cells in human peripheral blood induces increased expression of genes Wnt, Notch-1, BMI1 and HOXB4 through a signaling cascade PI3K/Akt/mTor/PTEN and promotes the formation of a self-renewing population of hematopoietic stem cells.[175]
Furthermore, dedifferentiation of progenitor cells induced by ACA-dependent signaling pathway leads to ACA-induced pluripotent stem cells, capable of differentiating in vitro into cells of all three germ layers.[176] The study of lectins' ability to maintain a culture of pluripotent human stem cells has led to the discovery of lectin Erythrina crista-galli (ECA), which can serve as a simple and highly effective matrix for the cultivation of human pluripotent stem cells.[177]
Reprogramming with a proteoglycan
An alternative strategy to convert somatic cells to pluripotent states may be continuous stimulation of fibroblasts by a single ECM proteoglycan, fibromodulin.[178] Such cells exhibit capability for skeletal muscle regeneration with markedly lower tumorigenic risk when compared to iPSCs.[179] The decreased tumorigenicity of such cells is related to CDKN2B upregulation during the recombinant human fibromodulin reprogramming process[180]
Reprogramming through a physical approach
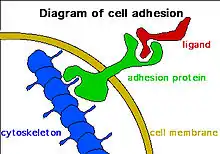
Cell adhesion protein E-cadherin is indispensable for a robust pluripotent phenotype.[181] During reprogramming for iPS cell generation, N-cadherin can replace function of E-cadherin.[182] These functions of cadherins are not directly related to adhesion because sphere morphology helps maintaining the "stemness" of stem cells.[183] Moreover, sphere formation, due to forced growth of cells on a low attachment surface, sometimes induces reprogramming. For example, neural progenitor cells can be generated from fibroblasts directly through a physical approach without introducing exogenous reprogramming factors.
Physical cues, in the form of parallel microgrooves on the surface of cell-adhesive substrates, can replace the effects of small-molecule epigenetic modifiers and significantly improve reprogramming efficiency. The mechanism relies on the mechanomodulation of the cells' epigenetic state. Specifically, "decreased histone deacetylase activity and upregulation of the expression of WD repeat domain 5 (WDR5) – a subunit of H3 methyltranferase – by microgrooved surfaces lead to increased histone H3 acetylation and methylation". Nanofibrous scaffolds with aligned fibre orientation produce effects similar to those produced by microgrooves, suggesting that changes in cell morphology may be responsible for modulation of the epigenetic state.[184]
Substrate rigidity is an important biophysical cue influencing neural induction and subtype specification. For example, soft substrates promote neuroepithelial conversion while inhibiting neural crest differentiation of hESCs in a BMP4-dependent manner. Mechanistic studies revealed a multi-targeted mechanotransductive process involving mechanosensitive Smad phosphorylation and nucleocytoplasmic shuttling, regulated by rigidity-dependent Hippo/YAP activities and actomyosin cytoskeleton integrity and contractility.[185]
Mouse embryonic stem cells (mESCs) undergo self-renewal in the presence of the cytokine leukemia inhibitory factor (LIF). Following LIF withdrawal, mESCs differentiate, accompanied by an increase in cell–substratum adhesion and cell spreading. Restricted cell spreading in the absence of LIF by either culturing mESCs on chemically defined, weakly adhesive biosubstrates, or by manipulating the cytoskeleton, allowed the cells to remain in an undifferentiated and pluripotent state. The effect of restricted cell spreading on mESC self-renewal is not mediated by increased intercellular adhesion, as inhibition of mESC adhesion using a function blocking anti E-cadherin antibody or siRNA does not promote differentiation.[186] Possible mechanisms of stem cell fate predetermination by physical interactions with the extracellular matrix have been described.[187][188]
A new method has been developed that turns cells into stem cells faster and more efficiently by 'squeezing' them using 3D microenvironment stiffness and density of the surrounding gel. The technique can be applied to a large number of cells to produce stem cells for medical purposes on an industrial scale.[189][190]
Cells involved in the reprogramming process change morphologically as the process proceeds. This results in physical differences in adhesive forces among cells. Substantial differences in 'adhesive signature' between pluripotent stem cells, partially reprogrammed cells, differentiated progeny and somatic cells allowed the development of separation process for isolation of pluripotent stem cells in microfluidic devices,[191] which is:
- fast (separation takes less than 10 minutes);
- efficient (separation results in a greater than 95 percent pure iPS cell culture);
- innocuous (cell survival rate is greater than 80 percent and the resulting cells retain normal transcriptional profiles, differentiation potential and karyotype).
Stem cells possess mechanical memory (they remember past physical signals) – with the Hippo signaling pathway factors:[192] Yes-associated protein (YAP) and transcriptional coactivator with PDZ-binding domain (TAZ) acting as an intracellular mechanical rheostat—that stores information from past physical environments and influences the cells' fate.[193][194]
Neural stem cells
Stroke and many neurodegenerative disorders, such as Parkinson's disease, Alzheimer's disease, and amyotrophic lateral sclerosis, need cell replacement therapies. The successful use of converted neural cells (cNs) in transplantations opens a new avenue to treat such diseases.[195] Nevertheless, induced neurons (iNs) directly converted from fibroblasts are terminally committed and exhibit very limited proliferative ability that may not provide enough autologous donor cells for transplantation.[196] Self-renewing induced neural stem cells (iNSCs) provide additional advantages over iNs for both basic research and clinical applications.[126][127][128][197][198]
For example, under specific growth conditions, mouse fibroblasts can be reprogrammed with a single factor, Sox2, to form iNSCs that self-renew in culture and after transplantation and can survive and integrate without forming tumors in mouse brains.[199] INSCs can be derived from adult human fibroblasts by non-viral techniques, thus offering a safe method for autologous transplantation or for the development of cell-based disease models.[198]
Neural chemically induced progenitor cells (ciNPCs) can be generated from mouse tail-tip fibroblasts and human urinary somatic cells without introducing exogenous factors, but - by a chemical cocktail, namely VCR (V, VPA, an inhibitor of HDACs; C, CHIR99021, an inhibitor of GSK-3 kinases and R, RepSox, an inhibitor of TGF beta signaling pathways), under a physiological hypoxic condition.[200] Alternative cocktails with inhibitors of histone deacetylation, glycogen synthase kinase and TGF-β pathways (where: sodium butyrate (NaB) or Trichostatin A (TSA) could replace VPA, Lithium chloride (LiCl) or lithium carbonate (Li2CO3) could substitute CHIR99021, or Repsox may be replaced with SB-431542 or Tranilast) show similar efficacies for ciNPC induction.[200] Zhang, et al.,[201] also report highly efficient reprogramming of mouse fibroblasts into induced neural stem cell-like cells (ciNSLCs) using a cocktail of nine components.
Multiple methods of direct transformation of somatic cells into induced neural stem cells have been described.[202]
Proof of principle experiments demonstrate that it is possible to convert transplanted human fibroblasts and human astrocytes directly in the brain that are engineered to express inducible forms of neural reprogramming genes, into neurons, when reprogramming genes (Ascl1, Brn2a and Myt1l) are activated after transplantation using a drug.[203]
Astrocytes – the most common neuroglial brain cells, which contribute to scar formation in response to injury – can be directly reprogrammed in vivo to become functional neurons that form networks in mice without the need for cell transplantation.[204] The researchers followed the mice for nearly a year to look for signs of tumor formation and reported finding none. The same researchers have turned scar-forming astrocytes into progenitor cells, called neuroblasts, that regenerated into neurons in the injured adult spinal cord.[205]
Oligodendrocyte precursor cells
Without myelin to insulate neurons, nerve signals quickly lose power. Diseases that attack myelin, such as multiple sclerosis, result in nerve signals that cannot propagate to nerve endings and as a consequence lead to cognitive, motor and sensory problems. Transplantation of oligodendrocyte precursor cells (OPCs), which can successfully create myelin sheaths around nerve cells, is a promising potential therapeutic response. Direct lineage conversion of mouse and rat fibroblasts into oligodendroglial cells provides a potential source of OPCs. Conversion by forced expression of both eight[206] or of the three[207] transcription factors Sox10, Olig2 and Zfp536, may provide such cells.
Cardiomyocytes
Cell-based in vivo therapies may provide a transformative approach to augment vascular and muscle growth and to prevent non-contractile scar formation by delivering transcription factors[123] or microRNAs[14] to the heart.[208] Cardiac fibroblasts, which represent 50% of the cells in the mammalian heart, can be reprogrammed into cardiomyocyte-like cells in vivo by local delivery of cardiac core transcription factors ( GATA4, MEF2C, TBX5 and for improved reprogramming plus ESRRG, MESP1, Myocardin and ZFPM2) after coronary ligation.[123][209] These results implicated therapies that can directly remuscularize the heart without cell transplantation. However, the efficiency of such reprogramming turned out to be very low and the phenotype of received cardiomyocyte-like cells does not resemble those of a mature normal cardiomyocyte. Furthermore, transplantation of cardiac transcription factors into injured murine hearts resulted in poor cell survival and minimal expression of cardiac genes.[210]
Meanwhile, advances in the methods of obtaining cardiac myocytes in vitro occurred.[211][212] Efficient cardiac differentiation of human iPS cells gave rise to progenitors that were retained within infarcted rat hearts and reduced remodeling of the heart after ischemic damage.[213]
The team of scientists, led by Sheng Ding, used a cocktail of nine chemicals (9C) for transdifferentiation of human skin cells into beating heart cells. With this method, more than 97% of the cells began beating, a characteristic of fully developed, healthy heart cells. The chemically induced cardiomyocyte-like cells (ciCMs) uniformly contracted and resembled human cardiomyocytes in their transcriptome, epigenetic, and electrophysiological properties. When transplanted into infarcted mouse hearts, 9C-treated fibroblasts were efficiently converted to ciCMs and developed into healthy-looking heart muscle cells within the organ.[214] This chemical reprogramming approach, after further optimization, may offer an easy way to provide the cues that induce heart muscle to regenerate locally.[215]
In another study, ischemic cardiomyopathy in the murine infarction model was targeted by iPS cell transplantation. It synchronized failing ventricles, offering a regenerative strategy to achieve resynchronization and protection from decompensation by dint of improved left ventricular conduction and contractility, reduced scarring and reversal of structural remodelling.[216] One protocol generated populations of up to 98% cardiomyocytes from hPSCs simply by modulating the canonical Wnt signaling pathway at defined time points in during differentiation, using readily accessible small molecule compounds.[217]
Discovery of the mechanisms controlling the formation of cardiomyocytes led to the development of the drug ITD-1, which effectively clears the cell surface from TGF-β receptor type II and selectively inhibits intracellular TGF-β signaling. It thus selectively enhances the differentiation of uncommitted mesoderm to cardiomyocytes, but not to vascular smooth muscle and endothelial cells.[218]
One project seeded decellularized mouse hearts with human iPSC-derived multipotential cardiovascular progenitor cells. The introduced cells migrated, proliferated and differentiated in situ into cardiomyocytes, smooth muscle cells and endothelial cells to reconstruct the hearts. In addition, the heart's extracellular matrix (the substrate of heart scaffold) signalled the human cells into becoming the specialised cells needed for proper heart function. After 20 days of perfusion with growth factors, the engineered heart tissues started to beat again and were responsive to drugs.[219]
Reprogramming of cardiac fibroblasts into induced cardiomyocyte-like cells (iCMs) in situ represents a promising strategy for cardiac regeneration. Mice exposed in vivo, to three cardiac transcription factors GMT (Gata4, Mef2c, Tbx5) and the small-molecules: SB-431542 (the transforming growth factor (TGF)-β inhibitor), and XAV939 (the WNT inhibitor) for 2 weeks after myocardial infarction showed significantly improved reprogramming (reprogramming efficiency increased eight-fold) and cardiac function compared to those exposed to only GMT.[220]
Rejuvenation of the muscle stem cell
The elderly often with progressive muscle weakness and regenerative failure owing in part to elevated activity of the p38α and p38β mitogen-activated kinase pathway in senescent skeletal muscle stem cells. Subjecting such stem cells to transient inhibition of p38α and p38β in conjunction with culture on soft hydrogel substrates rapidly expands and rejuvenates them that result in the return of their strength.[223]
In geriatric mice, resting satellite cells lose reversible quiescence by switching to an irreversible pre-senescence state, caused by derepression of p16INK4a (also called Cdkn2a). On injury, these cells fail to activate and expand, even in a youthful environment. p16INK4a silencing in geriatric satellite cells restores quiescence and muscle regenerative functions.[224]
Myogenic progenitors for potential use in disease modeling or cell-based therapies targeting skeletal muscle could also be generated directly from induced pluripotent stem cells using free-floating spherical culture (EZ spheres) in a culture medium supplemented with high concentrations (100 ng/ml) of fibroblast growth factor-2 (FGF-2) and epidermal growth factor.[225]
Hepatocytes
Unlike current protocols for deriving hepatocytes from human fibroblasts, Saiyong Zhu et al., (2014)[226] did not generate iPSCs but, using small molecules, cut short reprogramming to pluripotency to generate an induced multipotent progenitor cell (iMPC) state from which endoderm progenitor cells and subsequently hepatocytes (iMPC-Heps) were efficiently differentiated. After transplantation into an immune-deficient mouse model of human liver failure, iMPC-Heps proliferated extensively and acquired levels of hepatocyte function similar to those of human primary adult hepatocytes. iMPC-Heps did not form tumours, most probably because they never entered a pluripotent state.

These results establish the feasibility of significant liver repopulation of mice with human hepatocytes generated in vitro, which removes a long-standing roadblock on the path to autologous liver cell therapy.
A cocktail of small molecules, Y-27632, A-83-01 (a TGFβ kinase/activin receptor like kinase (ALK5) inhibitor), and CHIR99021 (potent inhibitor of GSK-3), can convert rat and mouse mature hepatocytes in vitro into proliferative bipotent cells – CLiPs (chemically induced liver progenitors). CLiPs can differentiate into both mature hepatocytes and biliary epithelial cells that can form functional ductal structures. In long-term culture CLiPs did not lose their proliferative capacity and their hepatic differentiation ability, and can repopulate chronically injured liver tissue.[227]
Insulin-producing cells
Complications of Diabetes mellitus such as cardiovascular diseases, retinopathy, neuropathy, nephropathy, and peripheral circulatory diseases depend on sugar dysregulation due to lack of insulin from pancreatic beta cells and can be lethal if they are not treated. One of the promising approaches to understand and cure diabetes is to use pluripotent stem cells (PSCs), including embryonic stem cells (ESCs) and induced PCSs (iPSCs).[228] Unfortunately, human PSC-derived insulin-expressing cells resemble human fetal β cells rather than adult β cells. In contrast to adult β cells, fetal β cells seem functionally immature, as indicated by increased basal glucose secretion and lack of glucose stimulation and confirmed by RNA-seq of whose transcripts.[229]
An alternative strategy is the conversion of fibroblasts towards distinct endodermal progenitor cell populations and, using cocktails of signalling factors, successful differentiation of these endodermal progenitor cells into functional beta-like cells both in vitro and in vivo.[230]
Overexpression of the three transcription factors, PDX1 (required for pancreatic bud outgrowth and beta-cell maturation), NGN3 (required for endocrine precursor cell formation) and MAFA (for beta-cell maturation) combination (called PNM) can lead to the transformation of some cell types into a beta cell-like state.[231] An accessible and abundant source of functional insulin-producing cells is intestine. PMN expression in human intestinal "organoids" stimulates the conversion of intestinal epithelial cells into β-like cells possibly acceptable for transplantation.[232]
Nephron Progenitors
Adult proximal tubule cells were directly transcriptionally reprogrammed to nephron progenitors of the embryonic kidney, using a pool of six genes of instructive transcription factors (SIX1, SIX2, OSR1, Eyes absent homolog 1(EYA1), Homeobox A11 (HOXA11) and Snail homolog 2 (SNAI2)) that activated genes consistent with a cap mesenchyme/nephron progenitor phenotype in the adult proximal tubule cell line.[233] The generation of such cells may lead to cellular therapies for adult renal disease. Embryonic kidney organoids placed into adult rat kidneys can undergo onward development and vascular development.[234]
Blood vessel cells
As blood vessels age, they often become abnormal in structure and function, thereby contributing to numerous age-associated diseases including myocardial infarction, ischemic stroke and atherosclerosis of arteries supplying the heart, brain and lower extremities. So, an important goal is to stimulate vascular growth for the collateral circulation to prevent the exacerbation of these diseases. Induced Vascular Progenitor Cells (iVPCs) are useful for cell-based therapy designed to stimulate coronary collateral growth. They were generated by partially reprogramming endothelial cells.[165] The vascular commitment of iVPCs is related to the epigenetic memory of endothelial cells, which engenders them as cellular components of growing blood vessels. That is why, when iVPCs were implanted into myocardium, they engrafted in blood vessels and increased coronary collateral flow better than iPSCs, mesenchymal stem cells, or native endothelial cells.[235]
Ex vivo genetic modification can be an effective strategy to enhance stem cell function. For example, cellular therapy employing genetic modification with Pim-1 kinase (a downstream effector of Akt, which positively regulates neovasculogenesis) of bone marrow–derived cells[236] or human cardiac progenitor cells, isolated from failing myocardium[237] results in durability of repair, together with the improvement of functional parameters of myocardial hemodynamic performance.
Stem cells extracted from fat tissue after liposuction may be coaxed into becoming progenitor smooth muscle cells (iPVSMCs) found in arteries and veins.[238]
The 2D culture system of human iPS cells[239] in conjunction with triple marker selection (CD34 (a surface glycophosphoprotein expressed on developmentally early embryonic fibroblasts), NP1 (receptor – neuropilin 1) and KDR (kinase insert domain-containing receptor)) for the isolation of vasculogenic precursor cells from human iPSC, generated endothelial cells that, after transplantation, formed stable, functional mouse blood vessels in vivo, lasting for 280 days.[240]
To treat infarction, it is important to prevent the formation of fibrotic scar tissue. This can be achieved in vivo by transient application of paracrine factors that redirect native heart progenitor stem cell contributions from scar tissue to cardiovascular tissue. For example, in a mouse myocardial infarction model, a single intramyocardial injection of human vascular endothelial growth factor A mRNA (VEGF-A modRNA), modified to escape the body's normal defense system, results in long-term improvement of heart function due to mobilization and redirection of epicardial progenitor cells toward cardiovascular cell types.[241]
Blood stem cells
Red blood cells
RBC transfusion is necessary for many patients. However, to date the supply of RBCs remains labile. In addition, transfusion risks infectious disease transmission. A large supply of safe RBCs generated in vitro would help to address this issue. Ex vivo erythroid cell generation may provide alternative transfusion products to meet present and future clinical requirements.[242][243] Red blood cells (RBC)s generated in vitro from mobilized CD34 positive cells have normal survival when transfused into an autologous recipient.[244] RBC produced in vitro contained exclusively fetal hemoglobin (HbF), which rescues the functionality of these RBCs. In vivo the switch of fetal to adult hemoglobin was observed after infusion of nucleated erythroid precursors derived from iPSCs.[245] Although RBCs do not have nuclei and therefore can not form a tumor, their immediate erythroblasts precursors have nuclei. The terminal maturation of erythroblasts into functional RBCs requires a complex remodeling process that ends with extrusion of the nucleus and the formation of an enucleated RBC.[246] Cell reprogramming often disrupts enucleation. Transfusion of in vitro-generated RBCs or erythroblasts does not sufficiently protect against tumor formation.
The aryl hydrocarbon receptor (AhR) pathway (which has been shown to be involved in the promotion of cancer cell development) plays an important role in normal blood cell development. AhR activation in human hematopoietic progenitor cells (HPs) drives an unprecedented expansion of HPs, megakaryocyte- and erythroid-lineage cells.[247] See also Concise Review:[248][249] The SH2B3 gene encodes a negative regulator of cytokine signaling and naturally occurring loss-of-function variants in this gene increase RBC counts in vivo. Targeted suppression of SH2B3 in primary human hematopoietic stem and progenitor cells enhanced the maturation and overall yield of in-vitro-derived RBCs. Moreover, inactivation of SH2B3 by CRISPR/Cas9 genome editing in human pluripotent stem cells allowed enhanced erythroid cell expansion with preserved differentiation.[250] (See also overview.[249][251])

Platelets
Platelets help prevent hemorrhage in thrombocytopenic patients and patients with thrombocythemia. A significant problem for multitransfused patients is refractoriness to platelet transfusions. Thus, the ability to generate platelet products ex vivo and platelet products lacking HLA antigens in serum-free media would have clinical value. An RNA interference-based mechanism used a lentiviral vector to express short-hairpin RNAi targeting β2-microglobulin transcripts in CD34-positive cells. Generated platelets demonstrated an 85% reduction in class I HLA antigens. These platelets appeared to have normal function in vitro[252][253]
One clinically applicable strategy for the derivation of functional platelets from human iPSC involves the establishment of stable immortalized megakaryocyte progenitor cell lines (imMKCLs) through doxycycline-dependent overexpression of BMI1 and BCL-XL. The resulting imMKCLs can be expanded in culture over extended periods (4–5 months), even after cryopreservation. Halting the overexpression (by the removal of doxycycline from the medium) of c-MYC, BMI1 and BCL-XL in growing imMKCLs led to the production of CD42b+ platelets with functionality comparable to that of native platelets on the basis of a range of assays in vitro and in vivo.[254] Thomas et al., describe a forward programming strategy relying on the concurrent exogenous expression of 3 transcription factors: GATA1, FLI1 and TAL1. The forward programmed megakaryocytes proliferate and differentiate in culture for several months with megakaryocyte purity over 90% reaching up to 2x105 mature megakaryocytes per input hPSC. Functional platelets are generated throughout the culture allowing the prospective collection of several transfusion units from as few as one million starting hPSCs.[255] See also overview[256]
Immune cells
A specialised type of white blood cell, known as cytotoxic T lymphocytes (CTLs), are produced by the immune system and are able to recognise specific markers on the surface of various infectious or tumour cells, causing them to launch an attack to kill the harmful cells. Thence, immunotherapy with functional antigen-specific T cells has potential as a therapeutic strategy for combating many cancers and viral infections.[257] However, cell sources are limited, because they are produced in small numbers naturally and have a short lifespan.
A potentially efficient approach for generating antigen-specific CTLs is to revert mature immune T cells into iPSCs, which possess indefinite proliferative capacity in vitro and after their multiplication to coax them to redifferentiate back into T cells.[258][259][260]
Another method combines iPSC and chimeric antigen receptor (CAR)[261] technologies to generate human T cells targeted to CD19, an antigen expressed by malignant B cells, in tissue culture.[262] This approach of generating therapeutic human T cells may be useful for cancer immunotherapy and other medical applications.
Invariant natural killer T (iNKT) cells have great clinical potential as adjuvants for cancer immunotherapy. iNKT cells act as innate T lymphocytes and serve as a bridge between the innate and acquired immune systems. They augment anti-tumor responses by producing interferon-gamma (IFN-γ).[263] The approach of collection, reprogramming/dedifferentiation, re-differentiation and injection has been proposed for related tumor treatment.[264]
Dendritic cells (DC) are specialized to control T-cell responses. DC with appropriate genetic modifications may survive long enough to stimulate antigen-specific CTL and after that be eliminated. DC-like antigen-presenting cells obtained from human induced pluripotent stem cells can serve as a source for vaccination therapy.[265]
CCAAT/enhancer binding protein-α (C/EBPα) induces transdifferentiation of B cells into macrophages at high efficiencies[266] and enhances reprogramming into iPS cells when co-expressed with transcription factors Oct4, Sox2, Klf4 and Myc.[267] with a 100-fold increase in iPS cell reprogramming efficiency, involving 95% of the population.[268] Furthermore, C/EBPa can convert selected human B cell lymphoma and leukemia cell lines into macrophage-like cells at high efficiencies, impairing the cells' tumor-forming capacity.[269]
Thymic epithelial cells rejuvenation
The thymus is the first organ to deteriorate as people age. This shrinking is one of the main reasons the immune system becomes less effective with age. Diminished expression of the thymic epithelial cell transcription factor FOXN1 has been implicated as a component of the mechanism regulating age-related involution.[270][271]
Clare Blackburn and colleagues show that established age-related thymic involution can be reversed by forced upregulation of just one transcription factor – FOXN1 in the thymic epithelial cells in order to promote rejuvenation, proliferation and differentiation of these cells into fully functional thymic epithelium.[272] This rejuvenation and increased proliferation was accompanied by upregulation of genes that promote cell cycle progression (cyclin D1, ΔNp63, FgfR2IIIb) and that are required in the thymic epithelial cells to promote specific aspects of T cell development (Dll4, Kitl, Ccl25, Cxcl12, Cd40, Cd80, Ctsl, Pax1). In the future, this method may be widely used to enhance immune function and combat Inflammaging in patients by rejuvenating the thymus in situ .[273]
Mesenchymal stem cells
Induction
Mesenchymal stem/stromal cells (MSCs) are under investigation for applications in cardiac, renal, neural, joint and bone repair, as well as in inflammatory conditions and hemopoietic cotransplantation.[274] This is because of their immunosuppressive properties and ability to differentiate into a wide range of mesenchymal-lineage tissues. MSCs are typically harvested from adult bone marrow or fat, but these require painful invasive procedures and are low-frequency sources, making up only 0.001–0.01% of bone marrow cells and 0.05% in liposuction aspirates.[275] Of concern for autologous use, in particular in the elderly most in need of tissue repair, MSCs decline in quantity and quality with age.[274][276][277]
IPSCs could be obtained by the cells rejuvenation of even centenarians.[9][41] Because iPSCs can be harvested free of ethical constraints and culture can be expanded indefinitely, they are an advantageous source of MSCs.[278] IPSC treatment with SB-431542 leads to rapid and uniform MSC generation from human iPSCs. (SB-431542 is an inhibitor of activin/TGF- pathways by blocking phosphorylation of ALK4, ALK5 and ALK7 receptors.) These iPS-MSCs may lack teratoma-forming ability, display a normal stable karyotype in culture and exhibit growth and differentiation characteristics that closely resemble those of primary MSCs. It has potential for in vitro scale-up, enabling MSC-based therapies.[279] MSC derived from iPSC have the capacity to aid periodontal regeneration and are a promising source of readily accessible stem cells for use in the clinical treatment of periodontitis.[280][281]
Lai et al., & Lu report the chemical method to generate MSC-like cells (iMSCs), from human primary dermal fibroblasts using six chemical inhibitors (SP600125, SB202190, Go6983, Y-27632, PD0325901, and CHIR99021) with or without 3 growth factors (transforming growth factor-β (TGF-β), basic fibroblast growth factor (bFGF), and leukemia inhibitory factor (LIF)). The chemical cocktail directly converts human fibroblasts to iMSCs with a monolayer culture in 6 days, and the conversion rate was approximately 38%.[282]
Besides cell therapy in vivo, the culture of human mesenchymal stem cells can be used in vitro for mass-production of exosomes, which are ideal vehicles for drug delivery.[283]
Dedifferentiated adipocytes
Adipose tissue, because of its abundance and relatively less invasive harvest methods, represents a source of mesenchymal stem cells (MSCs). Unfortunately, liposuction aspirates are only 0.05% MSCs.[275] However, a large amount of mature adipocytes, which in general have lost their proliferative abilities and therefore are typically discarded, can be easily isolated from the adipose cell suspension and dedifferentiated into lipid-free fibroblast-like cells, named dedifferentiated fat (DFAT) cells. DFAT cells re-establish active proliferation ability and express multipotent capacities.[284] Compared with adult stem cells, DFAT cells show unique advantages in abundance, isolation and homogeneity. Under proper induction culture in vitro or proper environment in vivo, DFAT cells could demonstrate adipogenic, osteogenic, chondrogenic and myogenic potentials. They could also exhibit perivascular characteristics and elicit neovascularization.[285][286][287]
Chondrogenic cells
Cartilage is the connective tissue responsible for frictionless joint movement. Its degeneration ultimately results in complete loss of joint function in the late stages of osteoarthritis. As an avascular and hypocellular tissue, cartilage has a limited capacity for self-repair. Chondrocytes are the only cell type in cartilage, in which they are surrounded by the extracellular matrix that they secrete and assemble.
One method of producing cartilage is to induce it from iPS cells.[288] Alternatively, it is possible to convert fibroblasts directly into induced chondrogenic cells (iChon) without an intermediate iPS cell stage, by inserting three reprogramming factors (c-MYC, KLF4 and SOX9).[289] Human iChon cells expressed marker genes for chondrocytes (type II collagen) but not fibroblasts.
Implanted into defects created in the articular cartilage of rats, human iChon cells survived to form cartilaginous tissue for at least four weeks, with no tumors. The method makes use of c-MYC, which is thought to have a major role in tumorigenesis and employs a retrovirus to introduce the reprogramming factors, excluding it from unmodified use in human therapy.[258][260][290]
Sources of cells for reprogramming
The most frequently used sources for reprogramming are blood cells[291][292][293][294][295] and fibroblasts, obtained by biopsy of the skin,[296] but taking cells from urine is less invasive.[297][298][299][300] The latter method does not require a biopsy or blood sampling. As of 2013, urine-derived stem cells had been differentiated into endothelial, osteogenic, chondrogenic, adipogenic, skeletal myogenic and neurogenic lineages, without forming teratomas.[301] Therefore, their epigenetic memory is suited to reprogramming into iPS cells. However, few cells appear in urine, only low conversion efficiencies had been achieved and the risk of bacterial contamination is relatively high.
Another promising source of cells for reprogramming are mesenchymal stem cells derived from human hair follicles.[302]
The origin of somatic cells used for reprogramming may influence the efficiency of reprogramming,[303][304] the functional properties of the resulting induced stem cells[305] and the ability to form tumors.[306][307]
IPSCs retain an epigenetic memory of their tissue of origin, which impacts their differentiation potential.[290][305][308][309][310][311] This epigenetic memory does not necessarily manifest itself at the pluripotency stage – iPSCs derived from different tissues exhibit proper morphology, express pluripotency markers and are able to differentiate into the three embryonic layers in vitro and in vivo. However, this epigenetic memory may manifest during re-differentiation into specific cell types that require the specific loci bearing residual epigenetic marks.
See also
- Transdifferentiation
- Examples of in vitro transdifferentiation by lineage-instructive approach
- Examples of in vitro transdifferentiation by initial epigenetic activation phase approach
- Examples of in vivo transdifferentiation by lineage-instructive approach
- Injury induced stem cell niches
- Transcription factors
- Growth factors
- Pioneer factors
- Cellular differentiation
- CAF-1
References
- ↑ Yamanaka S, Blau HM (June 2010). "Nuclear reprogramming to a pluripotent state by three approaches". Nature. 465 (7299): 704–12. Bibcode:2010Natur.465..704Y. doi:10.1038/nature09229. PMC 2901154. PMID 20535199.
- ↑ Gurdon JB, Wilmut I (June 2011). "Nuclear transfer to eggs and oocytes". Cold Spring Harbor Perspectives in Biology. 3 (6): a002659. doi:10.1101/cshperspect.a002659. PMC 3098674. PMID 21555407.
- 1 2 3 4 Tachibana M, Amato P, Sparman M, Gutierrez NM, Tippner-Hedges R, Ma H, et al. (June 2013). "Human embryonic stem cells derived by somatic cell nuclear transfer". Cell. 153 (6): 1228–38. doi:10.1016/j.cell.2013.05.006. PMC 3772789. PMID 23683578.
- ↑ Noggle S, Fung HL, Gore A, Martinez H, Satriani KC, Prosser R, et al. (October 2011). "Human oocytes reprogram somatic cells to a pluripotent state". Nature. 478 (7367): 70–5. Bibcode:2011Natur.478...70N. doi:10.1038/nature10397. PMID 21979046. S2CID 4370078.
- 1 2 Pan G, Wang T, Yao H, Pei D (June 2012). "Somatic cell reprogramming for regenerative medicine: SCNT vs. iPS cells". BioEssays. 34 (6): 472–6. doi:10.1002/bies.201100174. PMID 22419173. S2CID 205470867.
- ↑ Do JT, Han DW, Gentile L, Sobek-Klocke I, Stehling M, Lee HT, Schöler HR (April 2007). "Erasure of cellular memory by fusion with pluripotent cells". Stem Cells. 25 (4): 1013–20. doi:10.1634/stemcells.2006-0691. PMID 17218392.
- 1 2 Takahashi K, Tanabe K, Ohnuki M, Narita M, Ichisaka T, Tomoda K, Yamanaka S (November 2007). "Induction of pluripotent stem cells from adult human fibroblasts by defined factors". Cell. 131 (5): 861–72. doi:10.1016/j.cell.2007.11.019. hdl:2433/49782. PMID 18035408. S2CID 8531539.
- ↑ Wang W, Yang J, Liu H, Lu D, Chen X, Zenonos Z, et al. (November 2011). "Rapid and efficient reprogramming of somatic cells to induced pluripotent stem cells by retinoic acid receptor gamma and liver receptor homolog 1". Proceedings of the National Academy of Sciences of the United States of America. 108 (45): 18283–8. Bibcode:2011PNAS..10818283W. doi:10.1073/pnas.1100893108. PMC 3215025. PMID 21990348.
- 1 2 3 Lapasset L, Milhavet O, Prieur A, Besnard E, Babled A, Aït-Hamou N, et al. (November 2011). "Rejuvenating senescent and centenarian human cells by reprogramming through the pluripotent state". Genes & Development. 25 (21): 2248–53. doi:10.1101/gad.173922.111. PMC 3219229. PMID 22056670.
- ↑ Zhou H, Wu S, Joo JY, Zhu S, Han DW, Lin T, et al. (May 2009). "Generation of induced pluripotent stem cells using recombinant proteins". Cell Stem Cell. 4 (5): 381–4. doi:10.1016/j.stem.2009.04.005. PMC 10182564. PMID 19398399.
- ↑ Li Z, Rana TM (2012). "Using microRNAs to enhance the generation of induced pluripotent stem cells". Current Protocols in Stem Cell Biology. Vol. 20. pp. 4D.4.1–14. doi:10.1002/9780470151808.sc04a04s20. ISBN 978-0470151808. PMID 22415842.
- ↑ Anokye-Danso F, Trivedi CM, Juhr D, Gupta M, Cui Z, Tian Y, et al. (April 2011). "Highly efficient miRNA-mediated reprogramming of mouse and human somatic cells to pluripotency". Cell Stem Cell. 8 (4): 376–88. doi:10.1016/j.stem.2011.03.001. PMC 3090650. PMID 21474102.
- ↑ Miyoshi N, Ishii H, Nagano H, Haraguchi N, Dewi DL, Kano Y, et al. (June 2011). "Reprogramming of mouse and human cells to pluripotency using mature microRNAs". Cell Stem Cell. 8 (6): 633–8. doi:10.1016/j.stem.2011.05.001. PMID 21620789.
- 1 2 Jayawardena TM, Egemnazarov B, Finch EA, Zhang L, Payne JA, Pandya K, et al. (May 2012). "MicroRNA-mediated in vitro and in vivo direct reprogramming of cardiac fibroblasts to cardiomyocytes". Circulation Research. 110 (11): 1465–73. doi:10.1161/CIRCRESAHA.112.269035. PMC 3380624. PMID 22539765.
- ↑ Bao X, Zhu X, Liao B, Benda C, Zhuang Q, Pei D, et al. (April 2013). "MicroRNAs in somatic cell reprogramming". Current Opinion in Cell Biology. 25 (2): 208–14. doi:10.1016/j.ceb.2012.12.004. PMID 23332905.
- ↑ Yoshioka N, Gros E, Li HR, Kumar S, Deacon DC, Maron C, et al. (August 2013). "Efficient generation of human iPSCs by a synthetic self-replicative RNA". Cell Stem Cell. 13 (2): 246–54. doi:10.1016/j.stem.2013.06.001. PMC 3845961. PMID 23910086.
- 1 2 Hou P, Li Y, Zhang X, Liu C, Guan J, Li H, et al. (August 2013). "Pluripotent stem cells induced from mouse somatic cells by small-molecule compounds". Science. 341 (6146): 651–4. Bibcode:2013Sci...341..651H. doi:10.1126/science.1239278. PMID 23868920. S2CID 45685692.
Efe JA, Ding S (August 2011). "The evolving biology of small molecules: controlling cell fate and identity". Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 366 (1575): 2208–21. doi:10.1098/rstb.2011.0006. PMC 3130415. PMID 21727126. - 1 2 Stadtfeld M, Apostolou E, Ferrari F, Choi J, Walsh RM, Chen T, et al. (March 2012). "Ascorbic acid prevents loss of Dlk1-Dio3 imprinting and facilitates generation of all-iPS cell mice from terminally differentiated B cells". Nature Genetics. 44 (4): 398–405, S1-2. doi:10.1038/ng.1110. PMC 3538378. PMID 22387999.
- ↑ Pandian GN, Sugiyama H (June 2012). "Programmable genetic switches to control transcriptional machinery of pluripotency". Biotechnology Journal. 7 (6): 798–809. doi:10.1002/biot.201100361. PMID 22588775. S2CID 44394354.
Pandian GN, Nakano Y, Sato S, Morinaga H, Bando T, Nagase H, Sugiyama H (2012). "A synthetic small molecule for rapid induction of multiple pluripotency genes in mouse embryonic fibroblasts". Scientific Reports. 2: 544. Bibcode:2012NatSR...2E.544P. doi:10.1038/srep00544. PMC 3408130. PMID 22848790. - ↑ De Robertis EM (April 2006). "Spemann's organizer and self-regulation in amphibian embryos". Nature Reviews. Molecular Cell Biology. 7 (4): 296–302. doi:10.1038/nrm1855. PMC 2464568. PMID 16482093.
- ↑ Slack JM (January 2009). "Metaplasia and somatic cell reprogramming". The Journal of Pathology. 217 (2): 161–8. doi:10.1002/path.2442. PMID 18855879.
- ↑ Wei G, Schubiger G, Harder F, Müller AM (2000). "Stem cell plasticity in mammals and transdetermination in Drosophila: common themes?". Stem Cells. 18 (6): 409–14. doi:10.1634/stemcells.18-6-409. PMID 11072028.
Worley MI, Setiawan L, Hariharan IK (2012). "Regeneration and transdetermination in Drosophila imaginal discs". Annual Review of Genetics. 46: 289–310. doi:10.1146/annurev-genet-110711-155637. PMID 22934642. - ↑ Xu PF, Houssin N, Ferri-Lagneau KF, Thisse B, Thisse C (April 2014). "Construction of a vertebrate embryo from two opposing morphogen gradients". Science. 344 (6179): 87–9. Bibcode:2014Sci...344...87X. doi:10.1126/science.1248252. PMID 24700857. S2CID 43493203.
- ↑ Stange DE, Koo BK, Huch M, Sibbel G, Basak O, Lyubimova A, et al. (October 2013). "Differentiated Troy+ chief cells act as reserve stem cells to generate all lineages of the stomach epithelium". Cell. 155 (2): 357–68. doi:10.1016/j.cell.2013.09.008. PMC 4094146. PMID 24120136.
- ↑ Tata PR, Mou H, Pardo-Saganta A, Zhao R, Prabhu M, Law BM, et al. (November 2013). "Dedifferentiation of committed epithelial cells into stem cells in vivo". Nature. 503 (7475): 218–23. Bibcode:2013Natur.503..218T. doi:10.1038/nature12777. PMC 4035230. PMID 24196716.
- ↑ Kusaba T, Lalli M, Kramann R, Kobayashi A, Humphreys BD (January 2014). "Differentiated kidney epithelial cells repair injured proximal tubule". Proceedings of the National Academy of Sciences of the United States of America. 111 (4): 1527–32. Bibcode:2014PNAS..111.1527K. doi:10.1073/pnas.1310653110. PMC 3910580. PMID 24127583.
- 1 2 Sieweke MH, Allen JE (November 2013). "Beyond stem cells: self-renewal of differentiated macrophages". Science. 342 (6161): 1242974. doi:10.1126/science.1242974. PMID 24264994. S2CID 2742670.
- 1 2 Soucie EL, Weng Z, Geirsdóttir L, Molawi K, Maurizio J, Fenouil R, et al. (February 2016). "Lineage-specific enhancers activate self-renewal genes in macrophages and embryonic stem cells". Science. 351 (6274): aad5510. doi:10.1126/science.aad5510. PMC 4811353. PMID 26797145.
- ↑ Sandoval-Guzmán T, Wang H, Khattak S, Schuez M, Roensch K, Nacu E, et al. (February 2014). "Fundamental differences in dedifferentiation and stem cell recruitment during skeletal muscle regeneration in two salamander species". Cell Stem Cell. 14 (2): 174–87. doi:10.1016/j.stem.2013.11.007. PMID 24268695.
- ↑ Kuroda Y, Wakao S, Kitada M, Murakami T, Nojima M, Dezawa M (2013). "Isolation, culture and evaluation of multilineage-differentiating stress-enduring (Muse) cells". Nature Protocols. 8 (7): 1391–415. doi:10.1038/nprot.2013.076. PMID 23787896. S2CID 28597290.
- ↑ Kuroda Y, Kitada M, Wakao S, Nishikawa K, Tanimura Y, Makinoshima H, et al. (May 2010). "Unique multipotent cells in adult human mesenchymal cell populations". Proceedings of the National Academy of Sciences of the United States of America. 107 (19): 8639–43. Bibcode:2010PNAS..107.8639K. doi:10.1073/pnas.0911647107. PMC 2889306. PMID 20421459.
- ↑ Ogura F, Wakao S, Kuroda Y, Tsuchiyama K, Bagheri M, Heneidi S, et al. (April 2014). "Human adipose tissue possesses a unique population of pluripotent stem cells with nontumorigenic and low telomerase activities: potential implications in regenerative medicine". Stem Cells and Development. 23 (7): 717–28. doi:10.1089/scd.2013.0473. PMID 24256547.
- ↑ Heneidi S, Simerman AA, Keller E, Singh P, Li X, Dumesic DA, Chazenbalk G (2013). "Awakened by cellular stress: isolation and characterization of a novel population of pluripotent stem cells derived from human adipose tissue". PLOS ONE. 8 (6): e64752. Bibcode:2013PLoSO...864752H. doi:10.1371/journal.pone.0064752. PMC 3673968. PMID 23755141.
- ↑ Shigemoto T, Kuroda Y, Wakao S, Dezawa M (July 2013). "A novel approach to collecting satellite cells from adult skeletal muscles on the basis of their stress tolerance". Stem Cells Translational Medicine. 2 (7): 488–98. doi:10.5966/sctm.2012-0130. PMC 3697816. PMID 23748608.
- ↑ Sisakhtnezhad S, Matin MM (June 2012). "Transdifferentiation: a cell and molecular reprogramming process". Cell and Tissue Research. 348 (3): 379–96. doi:10.1007/s00441-012-1403-y. PMID 22526624. S2CID 6675120.
- ↑ Merrell AJ, Stanger BZ (July 2016). "Adult cell plasticity in vivo: de-differentiation and transdifferentiation are back in style". Nature Reviews. Molecular Cell Biology. 17 (7): 413–25. doi:10.1038/nrm.2016.24. PMC 5818993. PMID 26979497.
- ↑ Dinnyes A, Cindy Tian X, Oback B (17 April 2013). "Nuclear Transfer for Cloning Animals". In Meyers RA (ed.). Stem Cells: From Biology to Therapy. John Wiley & Sons. pp. 299–344. ISBN 978-3-527-66854-0.
- ↑ Jullien J, Pasque V, Halley-Stott RP, Miyamoto K, Gurdon JB (June 2011). "Mechanisms of nuclear reprogramming by eggs and oocytes: a deterministic process?". Nature Reviews. Molecular Cell Biology. 12 (7): 453–9. doi:10.1038/nrm3140. PMC 3657683. PMID 21697902.
- ↑ Campbell KH (March 2002). "A background to nuclear transfer and its applications in agriculture and human therapeutic medicine". Journal of Anatomy. 200 (Pt 3): 267–75. doi:10.1046/j.1469-7580.2002.00035.x. PMC 1570687. PMID 12033731.
- ↑ US 8,647,872 patent
- 1 2 Chung YG, Eum JH, Lee JE, Shim SH, Sepilian V, Hong SW, et al. (June 2014). "Human somatic cell nuclear transfer using adult cells". Cell Stem Cell. 14 (6): 777–80. doi:10.1016/j.stem.2014.03.015. PMID 24746675.
- ↑ Mahla RS (2016). "Stem Cells Applications in Regenerative Medicine and Disease Therapeutics". International Journal of Cell Biology. 2016 (7): 6940283. doi:10.1155/2016/6940283. PMC 4969512. PMID 27516776.
- ↑ Yang H, Shi L, Wang BA, Liang D, Zhong C, Liu W, et al. (April 2012). "Generation of genetically modified mice by oocyte injection of androgenetic haploid embryonic stem cells". Cell. 149 (3): 605–17. doi:10.1016/j.cell.2012.04.002. PMID 22541431.
- ↑ Hayashi K, Ogushi S, Kurimoto K, Shimamoto S, Ohta H, Saitou M (November 2012). "Offspring from oocytes derived from in vitro primordial germ cell-like cells in mice". Science. 338 (6109): 971–5. Bibcode:2012Sci...338..971H. doi:10.1126/science.1226889. PMID 23042295. S2CID 6196269.
- ↑ Kishigami S, Mizutani E, Ohta H, Hikichi T, Thuan NV, Wakayama S, et al. (February 2006). "Significant improvement of mouse cloning technique by treatment with trichostatin A after somatic nuclear transfer". Biochemical and Biophysical Research Communications. 340 (1): 183–9. doi:10.1016/j.bbrc.2005.11.164. PMID 16356478.
- ↑ Wakayama S, Kohda T, Obokata H, Tokoro M, Li C, Terashita Y, et al. (March 2013). "Successful serial recloning in the mouse over multiple generations". Cell Stem Cell. 12 (3): 293–7. doi:10.1016/j.stem.2013.01.005. PMID 23472871.
- ↑ "Official website of the Presidential Commission for the Study of Bioethical Issues". Archived from the original on 2008-09-16. Retrieved 2014-02-25.
- ↑ Paull D, Emmanuele V, Weiss KA, Treff N, Stewart L, Hua H, et al. (January 2013). "Nuclear genome transfer in human oocytes eliminates mitochondrial DNA variants". Nature. 493 (7434): 632–7. Bibcode:2013Natur.493..632P. doi:10.1038/nature11800. PMC 7924261. PMID 23254936.
- ↑ Tachibana M, Amato P, Sparman M, Woodward J, Sanchis DM, Ma H, et al. (January 2013). "Towards germline gene therapy of inherited mitochondrial diseases". Nature. 493 (7434): 627–31. Bibcode:2013Natur.493..627T. doi:10.1038/nature11647. PMC 3561483. PMID 23103867.
- ↑ Hayden EC (October 2013). "Regulators weigh benefits of 'three-parent' fertilization". Nature. 502 (7471): 284–5. Bibcode:2013Natur.502..284C. doi:10.1038/502284a. PMID 24132269.
- ↑ Le R, Kou Z, Jiang Y, Li M, Huang B, Liu W, et al. (January 2014). "Enhanced telomere rejuvenation in pluripotent cells reprogrammed via nuclear transfer relative to induced pluripotent stem cells". Cell Stem Cell. 14 (1): 27–39. doi:10.1016/j.stem.2013.11.005. PMID 24268696.
- ↑ Cibelli J, Lanza R, Campbell KH, West MD (14 September 2002). Principles of Cloning. Academic Press. ISBN 978-0-08-049215-5.
- ↑ Shinagawa T, Takagi T, Tsukamoto D, Tomaru C, Huynh LM, Sivaraman P, et al. (February 2014). "Histone variants enriched in oocytes enhance reprogramming to induced pluripotent stem cells". Cell Stem Cell. 14 (2): 217–27. doi:10.1016/j.stem.2013.12.015. PMID 24506885.
- 1 2 Abad M, Mosteiro L, Pantoja C, Cañamero M, Rayon T, Ors I, et al. (October 2013). "Reprogramming in vivo produces teratomas and iPS cells with totipotency features". Nature. 502 (7471): 340–5. Bibcode:2013Natur.502..340A. doi:10.1038/nature12586. PMID 24025773. S2CID 4448205.
Naik G (2013-09-11). "New Promise for Stem Cells - WSJ.com". Online.wsj.com. Retrieved 2014-01-30. - ↑ Choi YJ, Lin CP, Risso D, Chen S, Kim TA, Tan MH, et al. (February 2017). "Deficiency of microRNA miR-34a expands cell fate potential in pluripotent stem cells". Science. 355 (6325): eaag1927. doi:10.1126/science.aag1927. PMC 6138252. PMID 28082412.
- "miR-34a MicroRNA Deficiency Induces Totipotent Stem Cells". Sciencebeta. 2017-01-14.
- ↑ Hu, M., Wei, H., Zhang, J., Bai, Y., Gao, F., Li, L., & Zhang, S. (2013). Efficient production of chimeric mice from embryonic stem cells injected into 4-to 8-cell and blastocyst embryos. Journal of animal science and biotechnology, 4, 1-7. PMID 23514327 PMC 3622560 doi:10.1186/2049-1891-4-12
- ↑ Stevens LC (March 1970). "The development of transplantable teratocarcinomas from intratesticular grafts of pre- and postimplantation mouse embryos". Developmental Biology. 21 (3): 364–82. doi:10.1016/0012-1606(70)90130-2. PMID 5436899.
- ↑ Mintz B, Cronmiller C, Custer RP (June 1978). "Somatic cell origin of teratocarcinomas". Proceedings of the National Academy of Sciences of the United States of America. 75 (6): 2834–8. Bibcode:1978PNAS...75.2834M. doi:10.1073/pnas.75.6.2834. PMC 392659. PMID 275854.
- ↑ Mintz B, Illmensee K (September 1975). "Normal genetically mosaic mice produced from malignant teratocarcinoma cells". Proceedings of the National Academy of Sciences of the United States of America. 72 (9): 3585–9. Bibcode:1975PNAS...72.3585M. doi:10.1073/pnas.72.9.3585. PMC 433040. PMID 1059147.
- ↑ Martin GR, Evans MJ (April 1975). "Differentiation of clonal lines of teratocarcinoma cells: formation of embryoid bodies in vitro". Proceedings of the National Academy of Sciences of the United States of America. 72 (4): 1441–5. Bibcode:1975PNAS...72.1441M. doi:10.1073/pnas.72.4.1441. PMC 432551. PMID 1055416.
- ↑ Illmensee K, Mintz B (February 1976). "Totipotency and normal differentiation of single teratocarcinoma cells cloned by injection into blastocysts". Proceedings of the National Academy of Sciences of the United States of America. 73 (2): 549–53. Bibcode:1976PNAS...73..549I. doi:10.1073/pnas.73.2.549. PMC 335947. PMID 1061157.
- ↑ Martin GR (December 1981). "Isolation of a pluripotent cell line from early mouse embryos cultured in medium conditioned by teratocarcinoma stem cells". Proceedings of the National Academy of Sciences of the United States of America. 78 (12): 7634–8. Bibcode:1981PNAS...78.7634M. doi:10.1073/pnas.78.12.7634. PMC 349323. PMID 6950406.
- ↑ Martin GR (August 1980). "Teratocarcinomas and mammalian embryogenesis". Science. 209 (4458): 768–76. Bibcode:1980Sci...209..768M. doi:10.1126/science.6250214. PMID 6250214.
- ↑ Papaioannou VE, Gardner RL, McBurney MW, Babinet C, Evans MJ (April 1978). "Participation of cultured teratocarcinoma cells in mouse embryogenesis". Journal of Embryology and Experimental Morphology. 44: 93–104. PMID 650144.
- ↑ Graham CF (January 1977). "Teratocarcinoma cells and normal mouse embryogenesiseditor=Michael I. Sherman". Concepts in Mammalian Embryogenesis. MIT Press. ISBN 978-0-262-19158-6.
- ↑ Illmensee K (14 June 2012). "Reversion of Malignancy and Normalized Differentiation of Teratocarcinoma Cells in Chimeric Mice". In Russell LB (ed.). Genetic Mosaics and Chimeras in Mammals. Vol. 12. Springer London, Limited. pp. 3–24. doi:10.1007/978-1-4684-3390-6_1. ISBN 978-1-4684-3392-0. PMID 378217.
{{cite book}}:|journal=ignored (help) - ↑ Stuart HT, van Oosten AL, Radzisheuskaya A, Martello G, Miller A, Dietmann S, et al. (February 2014). "NANOG amplifies STAT3 activation and they synergistically induce the naive pluripotent program". Current Biology. 24 (3): 340–6. doi:10.1016/j.cub.2013.12.040. PMC 3982126. PMID 24462001.
- ↑ Choi J, Lee S, Mallard W, Clement K, Tagliazucchi GM, Lim H, et al. (November 2015). "A comparison of genetically matched cell lines reveals the equivalence of human iPSCs and ESCs". Nature Biotechnology. 33 (11): 1173–81. doi:10.1038/nbt.3388. PMC 4847940. PMID 26501951.
- ↑ Boland MJ, Hazen JL, Nazor KL, Rodriguez AR, Gifford W, Martin G, et al. (September 2009). "Adult mice generated from induced pluripotent stem cells". Nature. 461 (7260): 91–4. Bibcode:2009Natur.461...91B. doi:10.1038/nature08310. PMID 19672243. S2CID 4423755.
Kang L, Wang J, Zhang Y, Kou Z, Gao S (August 2009). "iPS cells can support full-term development of tetraploid blastocyst-complemented embryos". Cell Stem Cell. 5 (2): 135–8. doi:10.1016/j.stem.2009.07.001. PMID 19631602. - ↑ Velychko S, Adachi K, Kim KP, Hou Y, MacCarthy CM, Wu G, Schöler HR (December 2019). "Excluding Oct4 from Yamanaka Cocktail Unleashes the Developmental Potential of iPSCs". Cell Stem Cell. 25 (6): 737–753.e4. doi:10.1016/j.stem.2019.10.002. PMC 6900749. PMID 31708402.
- "Quality of induced pluripotent stem cells is dramatically enhanced by omitting what was thought to be the most crucial reprogramming factor". Max-Planck Institute. November 7, 2019.
- ↑ Yagi T, Kosakai A, Ito D, Okada Y, Akamatsu W, Nihei Y, et al. (2012). "Establishment of induced pluripotent stem cells from centenarians for neurodegenerative disease research". PLOS ONE. 7 (7): e41572. Bibcode:2012PLoSO...741572Y. doi:10.1371/journal.pone.0041572. PMC 3405135. PMID 22848530.
- ↑ Rohani L, Johnson AA, Arnold A, Stolzing A (February 2014). "The aging signature: a hallmark of induced pluripotent stem cells?". Aging Cell. 13 (1): 2–7. doi:10.1111/acel.12182. PMC 4326871. PMID 24256351.
- ↑ Yehezkel S, Rebibo-Sabbah A, Segev Y, Tzukerman M, Shaked R, Huber I, et al. (January 2011). "Reprogramming of telomeric regions during the generation of human induced pluripotent stem cells and subsequent differentiation into fibroblast-like derivatives". Epigenetics. 6 (1): 63–75. doi:10.4161/epi.6.1.13390. PMC 3052915. PMID 20861676.
- ↑ West MD, Vaziri H (July 2010). "Back to immortality: the restoration of embryonic telomere length during induced pluripotency". Regenerative Medicine. 5 (4): 485–8. doi:10.2217/rme.10.51. PMID 20632849.
- ↑ Marión RM, Blasco MA (April 2010). "Telomere rejuvenation during nuclear reprogramming". Current Opinion in Genetics & Development. 20 (2): 190–6. doi:10.1016/j.gde.2010.01.005. PMID 20176474.
Gourronc FA, Klingelhutz AJ (February 2012). "Therapeutic opportunities: telomere maintenance in inducible pluripotent stem cells". Mutation Research. 730 (1–2): 98–105. doi:10.1016/j.mrfmmm.2011.05.008. PMC 3179558. PMID 21605571. - ↑ Zhao T, Zhang ZN, Rong Z, Xu Y (May 2011). "Immunogenicity of induced pluripotent stem cells". Nature. 474 (7350): 212–5. doi:10.1038/nature10135. PMID 21572395. S2CID 4416964.
- ↑ Dhodapkar MV, Dhodapkar KM (March 2011). "Spontaneous and therapy-induced immunity to pluripotency genes in humans: clinical implications, opportunities and challenges". Cancer Immunology, Immunotherapy. 60 (3): 413–8. doi:10.1007/s00262-010-0944-8. PMC 3574640. PMID 21104412.
- ↑ Gutierrez-Aranda I, Ramos-Mejia V, Bueno C, Munoz-Lopez M, Real PJ, Mácia A, et al. (September 2010). "Human induced pluripotent stem cells develop teratoma more efficiently and faster than human embryonic stem cells regardless the site of injection". Stem Cells. 28 (9): 1568–70. doi:10.1002/stem.471. PMC 2996086. PMID 20641038.
- ↑ Chang CJ, Mitra K, Koya M, Velho M, Desprat R, Lenz J, Bouhassira EE (2011). "Production of embryonic and fetal-like red blood cells from human induced pluripotent stem cells". PLOS ONE. 6 (10): e25761. Bibcode:2011PLoSO...625761C. doi:10.1371/journal.pone.0025761. PMC 3192723. PMID 22022444.
- 1 2 Dabir DV, Hasson SA, Setoguchi K, Johnson ME, Wongkongkathep P, Douglas CJ, et al. (April 2013). "A small molecule inhibitor of redox-regulated protein translocation into mitochondria". Developmental Cell. 25 (1): 81–92. doi:10.1016/j.devcel.2013.03.006. PMC 3726224. PMID 23597483.
- ↑ Ben-David U, Gan QF, Golan-Lev T, Arora P, Yanuka O, Oren YS, et al. (February 2013). "Selective elimination of human pluripotent stem cells by an oleate synthesis inhibitor discovered in a high-throughput screen". Cell Stem Cell. 12 (2): 167–79. doi:10.1016/j.stem.2012.11.015. PMID 23318055.
- ↑ Lou KJ (2013). "Small molecules vs. Teratomas". Science-Business EXchange. 6 (7): 158. doi:10.1038/scibx.2013.158.
- ↑ Lee MO, Moon SH, Jeong HC, Yi JY, Lee TH, Shim SH, et al. (August 2013). "Inhibition of pluripotent stem cell-derived teratoma formation by small molecules". Proceedings of the National Academy of Sciences of the United States of America. 110 (35): E3281-90. Bibcode:2013PNAS..110E3281L. doi:10.1073/pnas.1303669110. PMC 3761568. PMID 23918355.
- ↑ Chaffer CL, Brueckmann I, Scheel C, Kaestli AJ, Wiggins PA, Rodrigues LO, et al. (May 2011). "Normal and neoplastic nonstem cells can spontaneously convert to a stem-like state". Proceedings of the National Academy of Sciences of the United States of America. 108 (19): 7950–5. Bibcode:2011PNAS..108.7950C. doi:10.1073/pnas.1102454108. PMC 3093533. PMID 21498687.
Fu W, Wang SJ, Zhou GD, Liu W, Cao Y, Zhang WJ (March 2012). "Residual undifferentiated cells during differentiation of induced pluripotent stem cells in vitro and in vivo". Stem Cells and Development. 21 (4): 521–9. doi:10.1089/scd.2011.0131. PMID 21631153. - ↑ Gurtan AM, Ravi A, Rahl PB, Bosson AD, JnBaptiste CK, Bhutkar A, et al. (April 2013). "Let-7 represses Nr6a1 and a mid-gestation developmental program in adult fibroblasts". Genes & Development. 27 (8): 941–54. doi:10.1101/gad.215376.113. PMC 3650230. PMID 23630078.
Wang H, Wang X, Xu X, Zwaka TP, Cooney AJ (December 2013). "Epigenetic reprogramming of the germ cell nuclear factor gene is required for proper differentiation of induced pluripotent cells". Stem Cells. 31 (12): 2659–66. doi:10.1002/stem.1367. PMC 3731425. PMID 23495137. - ↑ Lindgren AG, Natsuhara K, Tian E, Vincent JJ, Li X, Jiao J, et al. (January 2011). "Loss of Pten causes tumor initiation following differentiation of murine pluripotent stem cells due to failed repression of Nanog". PLOS ONE. 6 (1): e16478. Bibcode:2011PLoSO...616478L. doi:10.1371/journal.pone.0016478. PMC 3029365. PMID 21304588.
- ↑ Grad I, Hibaoui Y, Jaconi M, Chicha L, Bergström-Tengzelius R, Sailani MR, et al. (November 2011). "NANOG priming before full reprogramming may generate germ cell tumours". European Cells & Materials. 22: 258–74, discussio 274. doi:10.22203/ecm.v022a20. PMID 22071697.
- ↑ Okano H, Nakamura M, Yoshida K, Okada Y, Tsuji O, Nori S, et al. (February 2013). "Steps toward safe cell therapy using induced pluripotent stem cells". Circulation Research. 112 (3): 523–33. doi:10.1161/CIRCRESAHA.111.256149. PMID 23371901.
Cunningham JJ, Ulbright TM, Pera MF, Looijenga LH (September 2012). "Lessons from human teratomas to guide development of safe stem cell therapies". Nature Biotechnology. 30 (9): 849–57. doi:10.1038/nbt.2329. PMID 22965062. S2CID 20383770. - ↑ Bellin M, Marchetto MC, Gage FH, Mummery CL (November 2012). "Induced pluripotent stem cells: the new patient?". Nature Reviews. Molecular Cell Biology. 13 (11): 713–26. doi:10.1038/nrm3448. PMID 23034453. S2CID 37156639.
Sandoe J, Eggan K (July 2013). "Opportunities and challenges of pluripotent stem cell neurodegenerative disease models". Nature Neuroscience. 16 (7): 780–9. doi:10.1038/nn.3425. PMID 23799470. S2CID 9287796. - ↑ Takahashi K, Yamanaka S (June 2013). "Induced pluripotent stem cells in medicine and biology". Development. 140 (12): 2457–61. doi:10.1242/dev.092551. PMID 23715538.
Fu X, Xu Y (2012). "Challenges to the clinical application of pluripotent stem cells: towards genomic and functional stability". Genome Medicine. 4 (6): 55. doi:10.1186/gm354. PMC 3698533. PMID 22741526. - ↑ Araki R, Uda M, Hoki Y, Sunayama M, Nakamura M, Ando S, et al. (February 2013). "Negligible immunogenicity of terminally differentiated cells derived from induced pluripotent or embryonic stem cells". Nature. 494 (7435): 100–4. Bibcode:2013Natur.494..100A. doi:10.1038/nature11807. PMID 23302801. S2CID 205232231.
Wahlestedt M, Norddahl GL, Sten G, Ugale A, Frisk MA, Mattsson R, et al. (May 2013). "An epigenetic component of hematopoietic stem cell aging amenable to reprogramming into a young state". Blood. 121 (21): 4257–64. doi:10.1182/blood-2012-11-469080. PMID 23476050. S2CID 206916964. - ↑ Ohnishi K, Semi K, Yamamoto T, Shimizu M, Tanaka A, Mitsunaga K, et al. (February 2014). "Premature termination of reprogramming in vivo leads to cancer development through altered epigenetic regulation". Cell. 156 (4): 663–77. doi:10.1016/j.cell.2014.01.005. PMID 24529372.
- ↑ Browder KC, Reddy P, Yamamoto M, et al. (March 2022). "In vivo partial reprogramming alters age-associated molecular changes during physiological aging in mice". Nature Aging. 2 (3): 243–253. doi:10.1038/s43587-022-00183-2. PMID 37118377. S2CID 247305231.
- ↑ Ocampo A, Reddy P, Martinez-Redondo P, Platero-Luengo A, Hatanaka F, Hishida T, et al. (December 2016). "In Vivo Amelioration of Age-Associated Hallmarks by Partial Reprogramming". Cell. Elsevier BV. 167 (7): 1719–1733.e12. doi:10.1016/j.cell.2016.11.052. PMC 6300877. PMID 27984723.
- ↑ Olova N, Simpson DJ, Marioni RE, Chandra T (February 2019). "Partial reprogramming induces a steady decline in epigenetic age before loss of somatic identity". Aging Cell. Wiley. 18 (1): e12877. doi:10.1111/acel.12877. PMC 6351826. PMID 30450724.
- ↑ Chondronasiou D, Gill D, Mosteiro L, Urdinguio RG, Berenguer-Llergo A, Aguilera M, et al. (March 2022). "Multi-omic rejuvenation of naturally aged tissues by a single cycle of transient reprogramming". Aging Cell. 21 (3): e13578. doi:10.1111/acel.13578. PMC 8920440. PMID 35235716.
- ↑ Gill D, Parry A, Santos F, Okkenhaug H, Todd CD, Hernando-Herraez I, et al. (April 2022). "Multi-omic rejuvenation of human cells by maturation phase transient reprogramming". eLife. 11: e71624. bioRxiv 10.1101/2021.01.15.426786. doi:10.7554/eLife.71624. PMC 9023058. PMID 35390271.
- ↑ Lama SC (2022-04-17). "Skin cells made 30 years younger with new 'rejuvenation' technique". livescience.com. Retrieved 2022-04-24.
- ↑ Mfopou JK, De Groote V, Xu X, Heimberg H, Bouwens L (May 2007). "Sonic hedgehog and other soluble factors from differentiating embryoid bodies inhibit pancreas development". Stem Cells. 25 (5): 1156–65. doi:10.1634/stemcells.2006-0720. PMID 17272496.
- ↑ De Los Angeles A, Daley GQ (December 2013). "A chemical logic for reprogramming to pluripotency". Cell Research. 23 (12): 1337–8. doi:10.1038/cr.2013.119. PMC 3847567. PMID 23979017.
- ↑ Federation AJ, Bradner JE, Meissner A (March 2014). "The use of small molecules in somatic-cell reprogramming". Trends in Cell Biology. 24 (3): 179–87. doi:10.1016/j.tcb.2013.09.011. PMC 3943685. PMID 24183602.
- ↑ Zhao Y, Zhao T, Guan J, Zhang X, Fu Y, Ye J, et al. (December 2015). "A XEN-like State Bridges Somatic Cells to Pluripotency during Chemical Reprogramming". Cell. 163 (7): 1678–91. doi:10.1016/j.cell.2015.11.017. PMID 26686652.
- ↑ WO 2011071085, Hiromitsu N, Akihide K, Nao S, Keiichi I, Satoshi Y, "Method for Producing Cells Induced to Differentiate from Pluripotent Stem Cells", published 2011-06-16, assigned to University of Tokyo
- ↑ Amabile G, Welner RS, Nombela-Arrieta C, D'Alise AM, Di Ruscio A, Ebralidze AK, et al. (February 2013). "In vivo generation of transplantable human hematopoietic cells from induced pluripotent stem cells". Blood. 121 (8): 1255–64. doi:10.1182/blood-2012-06-434407. PMC 3701251. PMID 23212524.
- ↑ Suzuki N, Yamazaki S, Yamaguchi T, Okabe M, Masaki H, Takaki S, et al. (July 2013). "Generation of engraftable hematopoietic stem cells from induced pluripotent stem cells by way of teratoma formation". Molecular Therapy. 21 (7): 1424–31. doi:10.1038/mt.2013.71. PMC 3705943. PMID 23670574.
- ↑ Chou BK, Ye Z, Cheng L (July 2013). "Generation and homing of iPSC-derived hematopoietic cells in vivo". Molecular Therapy. 21 (7): 1292–3. doi:10.1038/mt.2013.129. PMC 3702102. PMID 23812546.
- ↑ Yamauchi T, Takenaka K, Urata S, Shima T, Kikushige Y, Tokuyama T, et al. (February 2013). "Polymorphic Sirpa is the genetic determinant for NOD-based mouse lines to achieve efficient human cell engraftment". Blood. 121 (8): 1316–25. doi:10.1182/blood-2012-06-440354. PMID 23293079.
- ↑ Rong Z, Wang M, Hu Z, Stradner M, Zhu S, Kong H, et al. (January 2014). "An effective approach to prevent immune rejection of human ESC-derived allografts". Cell Stem Cell. 14 (1): 121–30. doi:10.1016/j.stem.2013.11.014. PMC 4023958. PMID 24388175.
- ↑ Hirami Y, Osakada F, Takahashi K, Okita K, Yamanaka S, Ikeda H, et al. (July 2009). "Generation of retinal cells from mouse and human induced pluripotent stem cells". Neuroscience Letters. 458 (3): 126–31. doi:10.1016/j.neulet.2009.04.035. hdl:2433/97948. PMID 19379795. S2CID 10294931.
- ↑ Buchholz DE, Hikita ST, Rowland TJ, Friedrich AM, Hinman CR, Johnson LV, Clegg DO (October 2009). "Derivation of functional retinal pigmented epithelium from induced pluripotent stem cells". Stem Cells. 27 (10): 2427–34. doi:10.1002/stem.189. PMID 19658190.
- ↑ Yang J, Nong E, Tsang SH (2013). "Induced pluripotent stem cells and retinal degeneration treatment". Expert Rev. Ophthalmol. 8 (1): 5–8. doi:10.1586/EOP.12.75. S2CID 72809515.
- ↑ Fields MA, Hwang J, Gong J, Cai H, Del Priore L (9 December 2012). "The Eye as a Target Organ for Stem Cell Therapy". In Tsang S (ed.). Stem Cell Biology and Regenerative Medicine in Ophthalmology. Springer. pp. 1–30. ISBN 978-1-4614-5493-9.
- ↑ Li Y, Tsai YT, Hsu CW, Erol D, Yang J, Wu WH, et al. (December 2012). "Long-term safety and efficacy of human-induced pluripotent stem cell (iPS) grafts in a preclinical model of retinitis pigmentosa". Molecular Medicine. 18 (1): 1312–9. doi:10.2119/molmed.2012.00242. PMC 3521789. PMID 22895806.
- ↑ "Can stem cells make the blind see again?". Inquirer Lifestyle. July 15, 2013.
- ↑ Tzouvelekis A, Ntolios P, Bouros D (2013). "Stem cell treatment for chronic lung diseases". Respiration; International Review of Thoracic Diseases. 85 (3): 179–92. doi:10.1159/000346525. PMID 23364286.
- ↑ Wagner DE, Bonvillain RW, Jensen T, Girard ED, Bunnell BA, Finck CM, et al. (August 2013). "Can stem cells be used to generate new lungs? Ex vivo lung bioengineering with decellularized whole lung scaffolds". Respirology. 18 (6): 895–911. doi:10.1111/resp.12102. PMC 3729745. PMID 23614471.
- ↑ Wong AP, Rossant J (June 2013). "Generation of Lung Epithelium from Pluripotent Stem Cells". Current Pathobiology Reports. 1 (2): 137–145. doi:10.1007/s40139-013-0016-9. PMC 3646155. PMID 23662247.
- ↑ Mou H, Zhao R, Sherwood R, Ahfeldt T, Lapey A, Wain J, et al. (April 2012). "Generation of multipotent lung and airway progenitors from mouse ESCs and patient-specific cystic fibrosis iPSCs". Cell Stem Cell. 10 (4): 385–97. doi:10.1016/j.stem.2012.01.018. PMC 3474327. PMID 22482504.
- ↑ Ghaedi M, Calle EA, Mendez JJ, Gard AL, Balestrini J, Booth A, et al. (November 2013). "Human iPS cell-derived alveolar epithelium repopulates lung extracellular matrix". The Journal of Clinical Investigation. 123 (11): 4950–62. doi:10.1172/JCI68793. PMC 3809786. PMID 24135142.
- ↑ Ghaedi M, Mendez JJ, Bove PF, Sivarapatna A, Raredon MS, Niklason LE (January 2014). "Alveolar epithelial differentiation of human induced pluripotent stem cells in a rotating bioreactor". Biomaterials. 35 (2): 699–710. doi:10.1016/j.biomaterials.2013.10.018. PMC 3897000. PMID 24144903.
- ↑ Huang SX, Islam MN, O'Neill J, Hu Z, Yang YG, Chen YW, et al. (January 2014). "Efficient generation of lung and airway epithelial cells from human pluripotent stem cells". Nature Biotechnology. 32 (1): 84–91. doi:10.1038/nbt.2754. PMC 4101921. PMID 24291815.
- ↑ Niu Z, Hu Y, Chu Z, Yu M, Bai Y, Wang L, Hua J (January 2013). "Germ-like cell differentiation from induced pluripotent stem cells (iPSCs)". Cell Biochemistry and Function. 31 (1): 12–9. doi:10.1002/cbf.2924. PMID 23086862. S2CID 19237703.
Yang S, Bo J, Hu H, Guo X, Tian R, Sun C, et al. (April 2012). "Derivation of male germ cells from induced pluripotent stem cells in vitro and in reconstituted seminiferous tubules". Cell Proliferation. 45 (2): 91–100. doi:10.1111/j.1365-2184.2012.00811.x. PMC 6496715. PMID 22324506.
Panula S, Medrano JV, Kee K, Bergström R, Nguyen HN, Byers B, et al. (February 2011). "Human germ cell differentiation from fetal- and adult-derived induced pluripotent stem cells". Human Molecular Genetics. 20 (4): 752–62. doi:10.1093/hmg/ddq520. PMC 3024045. PMID 21131292. - 1 2 3 Qian L, Huang Y, Spencer CI, Foley A, Vedantham V, Liu L, et al. (May 2012). "In vivo reprogramming of murine cardiac fibroblasts into induced cardiomyocytes". Nature. 485 (7400): 593–8. Bibcode:2012Natur.485..593Q. doi:10.1038/nature11044. PMC 3369107. PMID 22522929.
- ↑ Szabo E, Rampalli S, Risueño RM, Schnerch A, Mitchell R, Fiebig-Comyn A, et al. (November 2010). "Direct conversion of human fibroblasts to multilineage blood progenitors". Nature. 468 (7323): 521–6. Bibcode:2010Natur.468..521S. doi:10.1038/nature09591. PMID 21057492. S2CID 205223078.
- ↑ Efe JA, Hilcove S, Kim J, Zhou H, Ouyang K, Wang G, et al. (March 2011). "Conversion of mouse fibroblasts into cardiomyocytes using a direct reprogramming strategy". Nature Cell Biology. 13 (3): 215–22. doi:10.1038/ncb2164. PMID 21278734. S2CID 5310172.
- 1 2 Lujan E, Chanda S, Ahlenius H, Südhof TC, Wernig M (February 2012). "Direct conversion of mouse fibroblasts to self-renewing, tripotent neural precursor cells". Proceedings of the National Academy of Sciences of the United States of America. 109 (7): 2527–32. Bibcode:2012PNAS..109.2527L. doi:10.1073/pnas.1121003109. PMC 3289376. PMID 22308465.
- 1 2 Thier M, Wörsdörfer P, Lakes YB, Gorris R, Herms S, Opitz T, et al. (April 2012). "Direct conversion of fibroblasts into stably expandable neural stem cells". Cell Stem Cell. 10 (4): 473–9. doi:10.1016/j.stem.2012.03.003. PMID 22445518.
- 1 2 3 Han DW, Tapia N, Hermann A, Hemmer K, Höing S, Araúzo-Bravo MJ, et al. (April 2012). "Direct reprogramming of fibroblasts into neural stem cells by defined factors". Cell Stem Cell. 10 (4): 465–72. doi:10.1016/j.stem.2012.02.021. PMID 22445517.
- ↑ Velychko S, Kang K, Kim SM, Kwak TH, Kim KP, Park C, et al. (April 2019). "Fusion of Reprogramming Factors Alters the Trajectory of Somatic Lineage Conversion". Cell Reports. 27 (1): 30–39.e4. doi:10.1016/j.celrep.2019.03.023. PMID 30943410.
- ↑ Taylor SM, Jones PA (August 1979). "Multiple new phenotypes induced in 10T1/2 and 3T3 cells treated with 5-azacytidine". Cell. 17 (4): 771–9. doi:10.1016/0092-8674(79)90317-9. PMID 90553. S2CID 45765608.
- ↑ Lassar AB, Paterson BM, Weintraub H (December 1986). "Transfection of a DNA locus that mediates the conversion of 10T1/2 fibroblasts to myoblasts". Cell. 47 (5): 649–56. doi:10.1016/0092-8674(86)90507-6. PMID 2430720. S2CID 46395399.
Davis RL, Weintraub H, Lassar AB (December 1987). "Expression of a single transfected cDNA converts fibroblasts to myoblasts". Cell. 51 (6): 987–1000. doi:10.1016/0092-8674(87)90585-x. PMID 3690668. S2CID 37741454.
Weintraub H, Tapscott SJ, Davis RL, Thayer MJ, Adam MA, Lassar AB, Miller AD (July 1989). "Activation of muscle-specific genes in pigment, nerve, fat, liver, and fibroblast cell lines by forced expression of MyoD". Proceedings of the National Academy of Sciences of the United States of America. 86 (14): 5434–8. Bibcode:1989PNAS...86.5434W. doi:10.1073/pnas.86.14.5434. PMC 297637. PMID 2748593. - ↑ Vierbuchen T, Wernig M (October 2011). "Direct lineage conversions: unnatural but useful?". Nature Biotechnology. 29 (10): 892–907. doi:10.1038/nbt.1946. PMC 3222779. PMID 21997635.
- ↑ Chandrakanthan V, Yeola A, Kwan JC, Oliver RA, Qiao Q, Kang YC, et al. (April 2016). "PDGF-AB and 5-Azacytidine induce conversion of somatic cells into tissue-regenerative multipotent stem cells". Proceedings of the National Academy of Sciences of the United States of America. 113 (16): E2306-15. Bibcode:2016PNAS..113E2306C. doi:10.1073/pnas.1518244113. PMC 4843430. PMID 27044077.
- ↑ Scientists develop 'game changing' stem cell repair system. Stem Cells Portal
- ↑ Could this new stem cell become the game changer for regenerative treatments? Archived 2016-04-07 at the Wayback Machine. Irish Examiner
- ↑ Riddle MR, Weintraub A, Nguyen KC, Hall DH, Rothman JH (December 2013). "Transdifferentiation and remodeling of post-embryonic C. elegans cells by a single transcription factor". Development. 140 (24): 4844–9. doi:10.1242/dev.103010. PMC 3848185. PMID 24257624.
- ↑ Wei S, Zou Q, Lai S, Zhang Q, Li L, Yan Q, et al. (January 2016). "Conversion of embryonic stem cells into extraembryonic lineages by CRISPR-mediated activators". Scientific Reports. 6: 19648. Bibcode:2016NatSR...619648W. doi:10.1038/srep19648. PMC 4726097. PMID 26782778.
- ↑ Black JB, Adler AF, Wang HG, D'Ippolito AM, Hutchinson HA, Reddy TE, et al. (September 2016). "Targeted Epigenetic Remodeling of Endogenous Loci by CRISPR/Cas9-Based Transcriptional Activators Directly Converts Fibroblasts to Neuronal Cells". Cell Stem Cell. 19 (3): 406–14. doi:10.1016/j.stem.2016.07.001. PMC 5010447. PMID 27524438.
- ↑ Jung DW, Williams DR (June 2011). "Novel chemically defined approach to produce multipotent cells from terminally differentiated tissue syncytia". ACS Chemical Biology. 6 (6): 553–62. doi:10.1021/cb2000154. PMID 21322636.
"In This Issue". ACS Chemical Biology. 7 (4): 619. 2012. doi:10.1021/cb300127f. - ↑ Zhang H, Wilson IA, Lerner RA (September 2012). "Selection of antibodies that regulate phenotype from intracellular combinatorial antibody libraries". Proceedings of the National Academy of Sciences of the United States of America. 109 (39): 15728–33. Bibcode:2012PNAS..10915728Z. doi:10.1073/pnas.1214275109. PMC 3465454. PMID 23019357.
- ↑ Xie J, Zhang H, Yea K, Lerner RA (May 2013). "Autocrine signaling based selection of combinatorial antibodies that transdifferentiate human stem cells". Proceedings of the National Academy of Sciences of the United States of America. 110 (20): 8099–104. Bibcode:2013PNAS..110.8099X. doi:10.1073/pnas.1306263110. PMC 3657831. PMID 23613575.
- ↑ Ring K (September 12, 2017). "CIRM-funded scientists discover a new way to make stem cells using antibodies".
- "Scripps Research Institute Scientists Find Antibody that Transforms Bone Marrow Stem Cells Directly into Brain Cells". The Scripts Research Institute. April 22, 2013. Archived from the original on 2014-02-01.
- ↑ Ito N, Ohta K (May 2015). "Reprogramming of human somatic cells by bacteria". Development, Growth & Differentiation. 57 (4): 305–12. doi:10.1111/dgd.12209. PMID 25866152.
- ↑ Ito N, Katoh K, Kushige H, Saito Y, Umemoto T, Matsuzaki Y, et al. (January 2018). "Ribosome Incorporation into Somatic Cells Promotes Lineage Transdifferentiation towards Multipotency". Scientific Reports. 8 (1): 1634. Bibcode:2018NatSR...8.1634I. doi:10.1038/s41598-018-20057-1. PMC 5786109. PMID 29374279.
- "Ribosomes Found to Induce Somatic Cell Pluripotency". Technology Networks. February 7, 2018.
- 1 2 3 4 Liu X, Ory V, Chapman S, Yuan H, Albanese C, Kallakury B, et al. (February 2012). "ROCK inhibitor and feeder cells induce the conditional reprogramming of epithelial cells". The American Journal of Pathology. 180 (2): 599–607. doi:10.1016/j.ajpath.2011.10.036. PMC 3349876. PMID 22189618.
- ↑ Rheinwald JG, Green H (November 1975). "Serial cultivation of strains of human epidermal keratinocytes: the formation of keratinizing colonies from single cells". Cell. 6 (3): 331–43. doi:10.1016/S0092-8674(75)80001-8. PMID 1052771. S2CID 53294766.
- ↑ Hiew YL (2011). Examining the biological consequences of DNA damage caused by irradiated J2-3T3 fibroblast feeder cells and HPV16: characterisation of the biological functions of Mll (PhD Thesis). University College London.
- ↑ Szumiel I (September 2012). "Radiation hormesis: Autophagy and other cellular mechanisms". International Journal of Radiation Biology. 88 (9): 619–28. doi:10.3109/09553002.2012.699698. PMID 22702489. S2CID 1472841.
- ↑ Kurosawa H (December 2012). "Application of Rho-associated protein kinase (ROCK) inhibitor to human pluripotent stem cells". Journal of Bioscience and Bioengineering. 114 (6): 577–81. doi:10.1016/j.jbiosc.2012.07.013. PMID 22898436.
- ↑ Terunuma A, Limgala RP, Park CJ, Choudhary I, Vogel JC (April 2010). "Efficient procurement of epithelial stem cells from human tissue specimens using a Rho-associated protein kinase inhibitor Y-27632". Tissue Engineering. Part A. 16 (4): 1363–8. doi:10.1089/ten.tea.2009.0339. PMC 2862604. PMID 19912046.
- ↑ Wu, X., Wang, S., Li, M., Li, J., Shen, J., Zhao, Y., ... & Wu, X. (2020). Conditional reprogramming: next generation cell culture. Acta Pharmaceutica Sinica B, 10(8), 1360-1381. PMID 32963937 PMC 7488362 doi:10.1016/j.apsb.2020.01.011
- ↑ Wu X, Wang S, Li M, Li J, Shen J, Zhao Y, et al. (August 2020). "Conditional reprogramming: next generation cell culture". Acta Pharmaceutica Sinica B. Elsevier BV. 10 (8): 1360–1381. doi:10.1016/j.apsb.2020.01.011. PMC 7488362. PMID 32963937.
- ↑ Chapman S, Liu X, Meyers C, Schlegel R, McBride AA (July 2010). "Human keratinocytes are efficiently immortalized by a Rho kinase inhibitor". The Journal of Clinical Investigation. 120 (7): 2619–26. doi:10.1172/JCI42297. PMC 2898606. PMID 20516646.
- ↑ Suprynowicz FA, Upadhyay G, Krawczyk E, Kramer SC, Hebert JD, Liu X, et al. (December 2012). "Conditionally reprogrammed cells represent a stem-like state of adult epithelial cells". Proceedings of the National Academy of Sciences of the United States of America. 109 (49): 20035–40. Bibcode:2012PNAS..10920035S. doi:10.1073/pnas.1213241109. PMC 3523865. PMID 23169653.
- 1 2 Agarwal S, Rimm DL (February 2012). "Making every cell like HeLa a giant step for cell culture". The American Journal of Pathology. 180 (2): 443–5. doi:10.1016/j.ajpath.2011.12.001. PMID 22192626.
- 1 2 Lisanti MP, Tanowitz HB (April 2012). "Translational discoveries, personalized medicine, and living biobanks of the future". The American Journal of Pathology. 180 (4): 1334–6. doi:10.1016/j.ajpath.2012.02.003. PMID 22453029.
- ↑ Li, D., Thomas, C., Shrivastava, N., Gersten, A., Gadsden, N., Schlecht, N., ... & Ow, T. J. (2023). Establishment of a diverse head and neck squamous cancer cell bank using conditional reprogramming culture methods. Journal of Medical Virology, 95(2), e28388. PMID 36477880 PMC 10168123 (available on 2024-02-01) doi:10.1002/jmv.28388
- ↑ Yuan H, Myers S, Wang J, Zhou D, Woo JA, Kallakury B, et al. (September 2012). "Use of reprogrammed cells to identify therapy for respiratory papillomatosis". The New England Journal of Medicine. 367 (13): 1220–7. doi:10.1056/NEJMoa1203055. PMC 4030597. PMID 23013073.
- ↑ Crystal AS, Shaw AT, Sequist LV, Friboulet L, Niederst MJ, Lockerman EL, et al. (December 2014). "Patient-derived models of acquired resistance can identify effective drug combinations for cancer". Science. 346 (6216): 1480–6. Bibcode:2014Sci...346.1480C. doi:10.1126/science.1254721. PMC 4388482. PMID 25394791.
- ↑ Palechor-Ceron N, Suprynowicz FA, Upadhyay G, Dakic A, Minas T, Simic V, et al. (December 2013). "Radiation induces diffusible feeder cell factor(s) that cooperate with ROCK inhibitor to conditionally reprogram and immortalize epithelial cells". The American Journal of Pathology. 183 (6): 1862–1870. doi:10.1016/j.ajpath.2013.08.009. PMC 5745544. PMID 24096078.
- ↑ Saenz FR, Ory V, AlOtaiby M, Rosenfield S, Furlong M, Cavalli LR, et al. (2014). "Conditionally reprogrammed normal and transformed mouse mammary epithelial cells display a progenitor-cell-like phenotype". PLOS ONE. 9 (5): e97666. Bibcode:2014PLoSO...997666S. doi:10.1371/journal.pone.0097666. PMC 4022745. PMID 24831228.
- ↑ Kaur S, Soto-Pantoja DR, Stein EV, Liu C, Elkahloun AG, Pendrak ML, et al. (2013). "Thrombospondin-1 signaling through CD47 inhibits self-renewal by regulating c-Myc and other stem cell transcription factors". Scientific Reports. 3: 1673. Bibcode:2013NatSR...3E1673K. doi:10.1038/srep01673. PMC 3628113. PMID 23591719.
- ↑ Soto-Pantoja DR, Ridnour LA, Wink DA, Roberts DD (2013). "Blockade of CD47 increases survival of mice exposed to lethal total body irradiation". Scientific Reports. 3: 1038. Bibcode:2013NatSR...3E1038S. doi:10.1038/srep01038. PMC 3539147. PMID 23301159.
- ↑ Gentek R, Molawi K, Sieweke MH (November 2014). "Tissue macrophage identity and self-renewal". Immunological Reviews. 262 (1): 56–73. doi:10.1111/imr.12224. PMID 25319327. S2CID 20849255.
- 1 2 Kurian L, Sancho-Martinez I, Nivet E, Aguirre A, Moon K, Pendaries C, et al. (January 2013). "Conversion of human fibroblasts to angioblast-like progenitor cells". Nature Methods. 10 (1): 77–83. doi:10.1038/nmeth.2255. PMC 3531579. PMID 23202434.
- ↑ Morris SA, Daley GQ (January 2013). "A blueprint for engineering cell fate: current technologies to reprogram cell identity". Cell Research. 23 (1): 33–48. doi:10.1038/cr.2013.1. PMC 3541663. PMID 23277278.
- ↑ Wang YC, Nakagawa M, Garitaonandia I, Slavin I, Altun G, Lacharite RM, et al. (November 2011). "Specific lectin biomarkers for isolation of human pluripotent stem cells identified through array-based glycomic analysis". Cell Research. 21 (11): 1551–63. doi:10.1038/cr.2011.148. PMC 3364725. PMID 21894191.
- ↑ Thomson JA, Itskovitz-Eldor J, Shapiro SS, Waknitz MA, Swiergiel JJ, Marshall VS, Jones JM (November 1998). "Embryonic stem cell lines derived from human blastocysts". Science. 282 (5391): 1145–7. Bibcode:1998Sci...282.1145T. doi:10.1126/science.282.5391.1145. PMID 9804556.
- ↑ Suila H, Hirvonen T, Ritamo I, Natunen S, Tuimala J, Laitinen S, et al. (April 2014). "Extracellular o-linked N-acetylglucosamine is enriched in stem cells derived from human umbilical cord blood". BioResearch Open Access. 3 (2): 39–44. doi:10.1089/biores.2013.0050. PMC 3995142. PMID 24804163.
- ↑ Perdigoto CN, Bardin AJ (February 2013). "Sending the right signal: Notch and stem cells". Biochimica et Biophysica Acta (BBA) - General Subjects. 1830 (2): 2307–22. doi:10.1016/j.bbagen.2012.08.009. PMID 22917651.
- ↑ Jafar-Nejad H, Leonardi J, Fernandez-Valdivia R (August 2010). "Role of glycans and glycosyltransferases in the regulation of Notch signaling". Glycobiology. 20 (8): 931–49. doi:10.1093/glycob/cwq053. PMC 2912550. PMID 20368670.
- ↑ Alisson-Silva F, de Carvalho Rodrigues D, Vairo L, Asensi KD, Vasconcelos-dos-Santos A, Mantuano NR, et al. (May 2014). "Evidences for the involvement of cell surface glycans in stem cell pluripotency and differentiation". Glycobiology. 24 (5): 458–68. doi:10.1093/glycob/cwu012. PMID 24578376.
- ↑ Hasehira K, Tateno H, Onuma Y, Ito Y, Asashima M, Hirabayashi J (December 2012). "Structural and quantitative evidence for dynamic glycome shift on production of induced pluripotent stem cells". Molecular & Cellular Proteomics. 11 (12): 1913–23. doi:10.1074/mcp.M112.020586. PMC 3518133. PMID 23023295.
- ↑ Becker Kojić ZA, Terness P (October 2002). "A novel human erythrocyte glycosylphosphatidylinositol (GPI)-anchored glycoprotein ACA. Isolation, purification, primary structure determination, and molecular parameters of its lipid structure". The Journal of Biological Chemistry. 277 (43): 40472–8. doi:10.1074/jbc.M202416200. PMID 12167612.
- ↑ Becker-Kojić ZA, Ureña-Peralta JR, Saffrich R, Rodriguez-Jiménez FJ, Rubio MP, Rios P, et al. (August 2013). "A novel human glycoprotein ACA is an upstream regulator of human hematopoiesis". Bulletin of Experimental Biology and Medicine. 155 (4): 536–51. doi:10.1007/s10517-013-2195-0. PMID 24143385. S2CID 15178878.
- ↑ Becker-Kojić ZA, Ureña-Peralta JR, Zipančić I, Rodriguez-Jiménez FJ, Rubio MP, Stojković P, et al. (August 2013). "Activation by ACA induces pluripotency in human blood progenitor cells". Bulletin of Experimental Biology and Medicine. 155 (4): 552–67. doi:10.1007/s10517-013-2196-z. PMID 24143386. S2CID 17978967.
- ↑ Mikkola M (2013). Human pluripotent stem cells: glycomic approaches for culturing and characterization. hdl:10138/37669. ISBN 978-952-10-8444-7.
- ↑ Zheng Z, Jian J, Zhang X, Zara JN, Yin W, Chiang M, et al. (August 2012). "Reprogramming of human fibroblasts into multipotent cells with a single ECM proteoglycan, fibromodulin". Biomaterials. Elsevier BV. 33 (24): 5821–31. doi:10.1016/j.biomaterials.2012.04.049. PMID 22622142. S2CID 206039904.
- ↑ Yang, P., Li, C., Lee, M., Marzvanyan, A., Zhao, Z. H., Ting, K., ... & Zheng, Z. (2020). Photopolymerizable hydrogel encapsulated fibromodulin-reprogrammed cells for muscle regeneration. Tissue engineering. Part A. https://doi.org/10.1089/ten.tea.2020.0026
- ↑ Zheng Z, Li C, Ha P, Chang GX, Yang P, Zhang X, et al. (July 2019). "CDKN2B upregulation prevents teratoma formation in multipotent fibromodulin-reprogrammed cells". The Journal of Clinical Investigation. American Society for Clinical Investigation. 129 (8): 3236–3251. doi:10.1172/jci125015. PMC 6668700. PMID 31305260.
- ↑ Redmer T, Diecke S, Grigoryan T, Quiroga-Negreira A, Birchmeier W, Besser D (July 2011). "E-cadherin is crucial for embryonic stem cell pluripotency and can replace OCT4 during somatic cell reprogramming". EMBO Reports. 12 (7): 720–6. doi:10.1038/embor.2011.88. PMC 3128971. PMID 21617704.
- ↑ Bedzhov I, Alotaibi H, Basilicata MF, Ahlborn K, Liszewska E, Brabletz T, Stemmler MP (November 2013). "Adhesion, but not a specific cadherin code, is indispensable for ES cell and induced pluripotency". Stem Cell Research. 11 (3): 1250–63. doi:10.1016/j.scr.2013.08.009. PMID 24036274.
- ↑ Su G, Zhao Y, Wei J, Xiao Z, Chen B, Han J, et al. (August 2013). "Direct conversion of fibroblasts into neural progenitor-like cells by forced growth into 3D spheres on low attachment surfaces". Biomaterials. 34 (24): 5897–906. doi:10.1016/j.biomaterials.2013.04.040. PMID 23680365.
- ↑ Downing TL, Soto J, Morez C, Houssin T, Fritz A, Yuan F, et al. (December 2013). "Biophysical regulation of epigenetic state and cell reprogramming". Nature Materials. 12 (12): 1154–62. Bibcode:2013NatMa..12.1154D. doi:10.1038/nmat3777. PMC 9675045. PMID 24141451.
- ↑ Sun Y, Yong KM, Villa-Diaz LG, Zhang X, Chen W, Philson R, et al. (June 2014). "Hippo/YAP-mediated rigidity-dependent motor neuron differentiation of human pluripotent stem cells". Nature Materials. 13 (6): 599–604. Bibcode:2014NatMa..13..599S. doi:10.1038/nmat3945. PMC 4051885. PMID 24728461.
- ↑ Murray P, Prewitz M, Hopp I, Wells N, Zhang H, Cooper A, et al. (November 2013). "The self-renewal of mouse embryonic stem cells is regulated by cell-substratum adhesion and cell spreading". The International Journal of Biochemistry & Cell Biology. 45 (11): 2698–705. doi:10.1016/j.biocel.2013.07.001. PMC 3898852. PMID 23871934.
- ↑ Guilak F, Cohen DM, Estes BT, Gimble JM, Liedtke W, Chen CS (July 2009). "Control of stem cell fate by physical interactions with the extracellular matrix". Cell Stem Cell. 5 (1): 17–26. doi:10.1016/j.stem.2009.06.016. PMC 2768283. PMID 19570510.
- ↑ Worley K, Certo A, Wan LQ (2012). "Geometry–Force Control of Stem Cell Fate". BioNanoScience. 3: 43–51. doi:10.1007/s12668-012-0067-0. S2CID 40888270.
- ↑ Caiazzo M, Okawa Y, Ranga A, Piersigilli A, Tabata Y, Lutolf MP (March 2016). "Defined three-dimensional microenvironments boost induction of pluripotency". Nature Materials. 15 (3): 344–52. Bibcode:2016NatMa..15..344C. doi:10.1038/nmat4536. hdl:1874/346253. PMID 26752655.
- "Squeezing cells into stem cells". ScienceDaily (Press release). January 11, 2016.
- ↑ Roy B, Yuan L, Lee Y, Bharti A, Mitra A, Shivashankar GV (May 2020). "Fibroblast rejuvenation by mechanical reprogramming and redifferentiation". Proceedings of the National Academy of Sciences of the United States of America. Proceedings of the National Academy of Sciences. 117 (19): 10131–10141. Bibcode:2020PNAS..11710131R. doi:10.1073/pnas.1911497117. PMC 7229653. PMID 32350144.
- ↑ Singh A, Suri S, Lee T, Chilton JM, Cooke MT, Chen W, et al. (May 2013). "Adhesion strength-based, label-free isolation of human pluripotent stem cells". Nature Methods. 10 (5): 438–44. doi:10.1038/nmeth.2437. PMC 3641175. PMID 23563795.
- ↑ Wang K, Degerny C, Xu M, Yang XJ (February 2009). "YAP, TAZ, and Yorkie: a conserved family of signal-responsive transcriptional coregulators in animal development and human disease". Biochemistry and Cell Biology. 87 (1): 77–91. doi:10.1139/O08-114. PMID 19234525.
- ↑ Yang C, Tibbitt MW, Basta L, Anseth KS (June 2014). "Mechanical memory and dosing influence stem cell fate". Nature Materials. 13 (6): 645–52. Bibcode:2014NatMa..13..645Y. doi:10.1038/nmat3889. PMC 4031270. PMID 24633344.
- ↑ Nampe D, Tsutsui H (December 2013). "Engineered micromechanical cues affecting human pluripotent stem cell regulations and fate". Journal of Laboratory Automation. 18 (6): 482–93. doi:10.1177/2211068213503156. PMID 24062363.
- ↑ Zhang W, Duan S, Li Y, Xu X, Qu J, Zhang W, Liu GH (February 2012). "Converted neural cells: induced to a cure?". Protein & Cell. 3 (2): 91–7. doi:10.1007/s13238-012-2029-2. PMC 4104580. PMID 22410787.
- ↑ Yang N, Ng YH, Pang ZP, Südhof TC, Wernig M (December 2011). "Induced neuronal cells: how to make and define a neuron". Cell Stem Cell. 9 (6): 517–25. doi:10.1016/j.stem.2011.11.015. PMC 4377331. PMID 22136927.
- ↑ Sheng C, Zheng Q, Wu J, Xu Z, Sang L, Wang L, et al. (April 2012). "Generation of dopaminergic neurons directly from mouse fibroblasts and fibroblast-derived neural progenitors". Cell Research. 22 (4): 769–72. doi:10.1038/cr.2012.32. PMC 3317566. PMID 22370632.
- 1 2 Maucksch C, Firmin E, Butler-Munro C, Montgomery J, Dottori M, Connor B (2012). "Non-Viral Generation of Neural Precursor-like Cells from Adult Human Fibroblasts". Journal of Stem Cells & Regenerative Medicine. 8 (3): 162–70. doi:10.46582/jsrm.0803009. PMC 3908292. PMID 24693194.
- ↑ Ring KL, Tong LM, Balestra ME, Javier R, Andrews-Zwilling Y, Li G, et al. (July 2012). "Direct reprogramming of mouse and human fibroblasts into multipotent neural stem cells with a single factor". Cell Stem Cell. 11 (1): 100–9. doi:10.1016/j.stem.2012.05.018. PMC 3399516. PMID 22683203.
- 1 2 Cheng L, Hu W, Qiu B, Zhao J, Yu Y, Guan W, et al. (June 2014). "Generation of neural progenitor cells by chemical cocktails and hypoxia". Cell Research. 24 (6): 665–79. doi:10.1038/cr.2014.32. PMC 4042166. PMID 24638034.
- ↑ Zhang M, Lin YH, Sun YJ, Zhu S, Zheng J, Liu K, et al. (May 2016). "Pharmacological Reprogramming of Fibroblasts into Neural Stem Cells by Signaling-Directed Transcriptional Activation". Cell Stem Cell. 18 (5): 653–67. doi:10.1016/j.stem.2016.03.020. PMC 4864020. PMID 27133794.
- ↑ Liu GH, Yi F, Suzuki K, Qu J, Izpisua Belmonte JC (July 2012). "Induced neural stem cells: a new tool for studying neural development and neurological disorders". Cell Research. 22 (7): 1087–91. doi:10.1038/cr.2012.73. PMC 3391017. PMID 22547025.
- ↑ Torper O, Pfisterer U, Wolf DA, Pereira M, Lau S, Jakobsson J, et al. (April 2013). "Generation of induced neurons via direct conversion in vivo". Proceedings of the National Academy of Sciences of the United States of America. 110 (17): 7038–43. Bibcode:2013PNAS..110.7038T. doi:10.1073/pnas.1303829110. PMC 3637783. PMID 23530235.
- ↑ Niu W, Zang T, Zou Y, Fang S, Smith DK, Bachoo R, Zhang CL (October 2013). "In vivo reprogramming of astrocytes to neuroblasts in the adult brain". Nature Cell Biology. 15 (10): 1164–75. doi:10.1038/ncb2843. PMC 3867822. PMID 24056302.
- ↑ Su Z, Niu W, Liu ML, Zou Y, Zhang CL (February 2014). "In vivo conversion of astrocytes to neurons in the injured adult spinal cord". Nature Communications. 5: 3338. Bibcode:2014NatCo...5.3338S. doi:10.1038/ncomms4338. PMC 3966078. PMID 24569435.
- ↑ Najm FJ, Lager AM, Zaremba A, Wyatt K, Caprariello AV, Factor DC, et al. (May 2013). "Transcription factor-mediated reprogramming of fibroblasts to expandable, myelinogenic oligodendrocyte progenitor cells". Nature Biotechnology. 31 (5): 426–33. doi:10.1038/nbt.2561. PMC 3678540. PMID 23584611.
- ↑ Yang N, Zuchero JB, Ahlenius H, Marro S, Ng YH, Vierbuchen T, et al. (May 2013). "Generation of oligodendroglial cells by direct lineage conversion". Nature Biotechnology. 31 (5): 434–9. doi:10.1038/nbt.2564. PMC 3677690. PMID 23584610.
- ↑ Xu C (October 2012). "Turning cardiac fibroblasts into cardiomyocytes in vivo". Trends in Molecular Medicine. 18 (10): 575–6. doi:10.1016/j.molmed.2012.06.009. PMID 22770847.
- ↑ Fu JD, Stone NR, Liu L, Spencer CI, Qian L, Hayashi Y, et al. (2013). "Direct reprogramming of human fibroblasts toward a cardiomyocyte-like state". Stem Cell Reports. 1 (3): 235–47. doi:10.1016/j.stemcr.2013.07.005. PMC 3849259. PMID 24319660.
- ↑ Chen JX, Krane M, Deutsch MA, Wang L, Rav-Acha M, Gregoire S, et al. (June 2012). "Inefficient reprogramming of fibroblasts into cardiomyocytes using Gata4, Mef2c, and Tbx5". Circulation Research. 111 (1): 50–5. doi:10.1161/CIRCRESAHA.112.270264. PMC 3390172. PMID 22581928.
- ↑ Burridge PW, Keller G, Gold JD, Wu JC (January 2012). "Production of de novo cardiomyocytes: human pluripotent stem cell differentiation and direct reprogramming". Cell Stem Cell. 10 (1): 16–28. doi:10.1016/j.stem.2011.12.013. PMC 3255078. PMID 22226352.
- ↑ Wang H, Cao N, Spencer CI, Nie B, Ma T, Xu T, et al. (March 2014). "Small molecules enable cardiac reprogramming of mouse fibroblasts with a single factor, Oct4". Cell Reports. 6 (5): 951–60. doi:10.1016/j.celrep.2014.01.038. PMC 4004339. PMID 24561253.
- ↑ Carpenter L, Carr C, Yang CT, Stuckey DJ, Clarke K, Watt SM (April 2012). "Efficient differentiation of human induced pluripotent stem cells generates cardiac cells that provide protection following myocardial infarction in the rat". Stem Cells and Development. 21 (6): 977–86. doi:10.1089/scd.2011.0075. PMC 3315757. PMID 22182484.
- ↑ Cao N, Huang Y, Zheng J, Spencer CI, Zhang Y, Fu JD, et al. (June 2016). "Conversion of human fibroblasts into functional cardiomyocytes by small molecules". Science. 352 (6290): 1216–20. Bibcode:2016Sci...352.1216C. doi:10.1126/science.aaf1502. PMID 27127239.
- ↑ Scientists Turn Skin Cells into Heart Cells and Brain Cells Using Drugs. Gladstone Institutes. News Center
- ↑ Yamada S, Nelson TJ, Kane GC, Martinez-Fernandez A, Crespo-Diaz RJ, Ikeda Y, et al. (September 2013). "Induced pluripotent stem cell intervention rescues ventricular wall motion disparity, achieving biological cardiac resynchronization post-infarction". The Journal of Physiology. 591 (17): 4335–49. doi:10.1113/jphysiol.2013.252288. PMC 3779120. PMID 23568891.
- ↑ Lian X, Hsiao C, Wilson G, Zhu K, Hazeltine LB, Azarin SM, et al. (July 2012). "Robust cardiomyocyte differentiation from human pluripotent stem cells via temporal modulation of canonical Wnt signaling". Proceedings of the National Academy of Sciences of the United States of America. 109 (27): E1848-57. doi:10.1073/pnas.1200250109. PMC 3390875. PMID 22645348.
- ↑ Willems E, Cabral-Teixeira J, Schade D, Cai W, Reeves P, Bushway PJ, et al. (August 2012). "Small molecule-mediated TGF-β type II receptor degradation promotes cardiomyogenesis in embryonic stem cells". Cell Stem Cell. 11 (2): 242–52. doi:10.1016/j.stem.2012.04.025. PMC 3419596. PMID 22862949.
- ↑ Lu TY, Lin B, Kim J, Sullivan M, Tobita K, Salama G, Yang L (2013). "Repopulation of decellularized mouse heart with human induced pluripotent stem cell-derived cardiovascular progenitor cells". Nature Communications. 4: 2307. Bibcode:2013NatCo...4.2307L. doi:10.1038/ncomms3307. PMID 23942048.
- ↑ Mohamed TM, Stone NR, Berry EC, Radzinsky E, Huang Y, Pratt K, et al. (March 2017). "Chemical Enhancement of In Vitro and In Vivo Direct Cardiac Reprogramming". Circulation. 135 (10): 978–995. doi:10.1161/CIRCULATIONAHA.116.024692. PMC 5340593. PMID 27834668.
- ↑ Budniatzky I, Gepstein L (April 2014). "Concise review: reprogramming strategies for cardiovascular regenerative medicine: from induced pluripotent stem cells to direct reprogramming". Stem Cells Translational Medicine. 3 (4): 448–57. doi:10.5966/sctm.2013-0163. PMC 3973716. PMID 24591731. Archived from the original on 2014-04-07.
- ↑ Funakoshi, S., Yoshida, Y. (2021). Recent progress of iPSC technology in cardiac diseases. Arch Toxicol 95, 3633–3650 doi:10.1007/s00204-021-03172-3
- ↑ Cosgrove BD, Gilbert PM, Porpiglia E, Mourkioti F, Lee SP, Corbel SY, et al. (March 2014). "Rejuvenation of the muscle stem cell population restores strength to injured aged muscles". Nature Medicine. 20 (3): 255–64. doi:10.1038/nm.3464. PMC 3949152. PMID 24531378.
- ↑ Sousa-Victor P, Gutarra S, García-Prat L, Rodriguez-Ubreva J, Ortet L, Ruiz-Bonilla V, et al. (February 2014). "Geriatric muscle stem cells switch reversible quiescence into senescence". Nature. 506 (7488): 316–21. Bibcode:2014Natur.506..316S. doi:10.1038/nature13013. PMID 24522534. S2CID 205237468.
- ↑ Hosoyama T, McGivern JV, Van Dyke JM, Ebert AD, Suzuki M (May 2014). "Derivation of myogenic progenitors directly from human pluripotent stem cells using a sphere-based culture". Stem Cells Translational Medicine. 3 (5): 564–74. doi:10.5966/sctm.2013-0143. PMC 4006483. PMID 24657962.
- ↑ Zhu S, Rezvani M, Harbell J, Mattis AN, Wolfe AR, Benet LZ, et al. (April 2014). "Mouse liver repopulation with hepatocytes generated from human fibroblasts". Nature. 508 (7494): 93–7. Bibcode:2014Natur.508...93Z. doi:10.1038/nature13020. PMC 4161230. PMID 24572354.
- ↑ Katsuda T, Kawamata M, Hagiwara K, Takahashi RU, Yamamoto Y, Camargo FD, Ochiya T (January 2017). "Conversion of Terminally Committed Hepatocytes to Culturable Bipotent Progenitor Cells with Regenerative Capacity". Cell Stem Cell. 20 (1): 41–55. doi:10.1016/j.stem.2016.10.007. PMID 27840021.
- ↑ Abdelalim EM, Bonnefond A, Bennaceur-Griscelli A, Froguel P (June 2014). "Pluripotent stem cells as a potential tool for disease modelling and cell therapy in diabetes". Stem Cell Reviews and Reports. 10 (3): 327–37. doi:10.1007/s12015-014-9503-6. PMID 24577791. S2CID 14364185.
- ↑ Hrvatin S, O'Donnell CW, Deng F, Millman JR, Pagliuca FW, DiIorio P, et al. (February 2014). "Differentiated human stem cells resemble fetal, not adult, β cells". Proceedings of the National Academy of Sciences of the United States of America. 111 (8): 3038–43. Bibcode:2014PNAS..111.3038H. doi:10.1073/pnas.1400709111. PMC 3939927. PMID 24516164.
- ↑ Zhu S, Russ HA, Wang X, Zhang M, Ma T, Xu T, et al. (January 2016). "Human pancreatic beta-like cells converted from fibroblasts". Nature Communications. 7: 10080. Bibcode:2016NatCo...710080Z. doi:10.1038/ncomms10080. PMC 4729817. PMID 26733021.
- ↑ Akinci E, Banga A, Tungatt K, Segal J, Eberhard D, Dutton JR, Slack JM (2013). "Reprogramming of various cell types to a beta-like state by Pdx1, Ngn3 and MafA". PLOS ONE. 8 (11): e82424. Bibcode:2013PLoSO...882424A. doi:10.1371/journal.pone.0082424. PMC 3843737. PMID 24312421.
- ↑ Chen YJ, Finkbeiner SR, Weinblatt D, Emmett MJ, Tameire F, Yousefi M, et al. (March 2014). "De novo formation of insulin-producing "neo-β cell islets" from intestinal crypts". Cell Reports. 6 (6): 1046–1058. doi:10.1016/j.celrep.2014.02.013. PMC 4245054. PMID 24613355.
- ↑ Hendry CE, Vanslambrouck JM, Ineson J, Suhaimi N, Takasato M, Rae F, Little MH (September 2013). "Direct transcriptional reprogramming of adult cells to embryonic nephron progenitors". Journal of the American Society of Nephrology. 24 (9): 1424–34. doi:10.1681/ASN.2012121143. PMC 3752949. PMID 23766537.
- ↑ Xinaris C, Benedetti V, Rizzo P, Abbate M, Corna D, Azzollini N, et al. (November 2012). "In vivo maturation of functional renal organoids formed from embryonic cell suspensions". Journal of the American Society of Nephrology. 23 (11): 1857–68. doi:10.1681/ASN.2012050505. PMC 3482737. PMID 23085631.
- ↑ Yin L, Ohanyan V, Pung YF, Delucia A, Bailey E, Enrick M, et al. (January 2012). "Induction of vascular progenitor cells from endothelial cells stimulates coronary collateral growth". Circulation Research. 110 (2): 241–52. doi:10.1161/CIRCRESAHA.111.250126. PMC 3974272. PMID 22095729.
- ↑ Quijada P, Toko H, Fischer KM, Bailey B, Reilly P, Hunt KD, et al. (June 2012). "Preservation of myocardial structure is enhanced by pim-1 engineering of bone marrow cells". Circulation Research. 111 (1): 77–86. doi:10.1161/CIRCRESAHA.112.265207. PMC 3398618. PMID 22619278.
- ↑ Mohsin S, Khan M, Toko H, Bailey B, Cottage CT, Wallach K, et al. (October 2012). "Human cardiac progenitor cells engineered with Pim-I kinase enhance myocardial repair". Journal of the American College of Cardiology. 60 (14): 1278–87. doi:10.1016/j.jacc.2012.04.047. PMC 3461098. PMID 22841153.
- ↑ Brennan JA, Arrizabalaga JH, Nollert MU (4 April 2018). "Development of a Human Tissue-Engineered Blood Vessel from Adipose-Derived Stem Cells". Circulation Research. 111: A360. doi:10.1161/res.111.suppl_1.a360.
- "Adult stem cells from liposuction used to create blood vessels in the lab". ScienceDaily (Press release). July 25, 2012.
- ↑ Wang ZZ, Au P, Chen T, Shao Y, Daheron LM, Bai H, et al. (March 2007). "Endothelial cells derived from human embryonic stem cells form durable blood vessels in vivo". Nature Biotechnology. 25 (3): 317–8. doi:10.1038/nbt1287. PMID 17322871. S2CID 25295979.
- ↑ Samuel R, Daheron L, Liao S, Vardam T, Kamoun WS, Batista A, et al. (July 2013). "Generation of functionally competent and durable engineered blood vessels from human induced pluripotent stem cells". Proceedings of the National Academy of Sciences of the United States of America. 110 (31): 12774–9. Bibcode:2013PNAS..11012774S. doi:10.1073/pnas.1310675110. PMC 3732948. PMID 23861493.
- ↑ Zangi L, Lui KO, von Gise A, Ma Q, Ebina W, Ptaszek LM, et al. (October 2013). "Modified mRNA directs the fate of heart progenitor cells and induces vascular regeneration after myocardial infarction". Nature Biotechnology. 31 (10): 898–907. doi:10.1038/nbt.2682. PMC 4058317. PMID 24013197.
- ↑ Zeuner A, Martelli F, Vaglio S, Federici G, Whitsett C, Migliaccio AR (August 2012). "Concise review: stem cell-derived erythrocytes as upcoming players in blood transfusion". Stem Cells. 30 (8): 1587–96. doi:10.1002/stem.1136. PMC 3697769. PMID 22644674.
- ↑ Hirose S, Takayama N, Nakamura S, Nagasawa K, Ochi K, Hirata S, et al. (2013). "Immortalization of erythroblasts by c-MYC and BCL-XL enables large-scale erythrocyte production from human pluripotent stem cells". Stem Cell Reports. 1 (6): 499–508. doi:10.1016/j.stemcr.2013.10.010. PMC 3871399. PMID 24371805.
- ↑ Giarratana MC, Rouard H, Dumont A, Kiger L, Safeukui I, Le Pennec PY, et al. (November 2011). "Proof of principle for transfusion of in vitro-generated red blood cells". Blood. 118 (19): 5071–9. doi:10.1182/blood-2011-06-362038. PMC 3217398. PMID 21885599.
- ↑ Kobari L, Yates F, Oudrhiri N, Francina A, Kiger L, Mazurier C, et al. (December 2012). "Human induced pluripotent stem cells can reach complete terminal maturation: in vivo and in vitro evidence in the erythropoietic differentiation model". Haematologica. 97 (12): 1795–803. doi:10.3324/haematol.2011.055566. PMC 3590085. PMID 22733021.
- ↑ Keerthivasan G, Wickrema A, Crispino JD (2011). "Erythroblast enucleation". Stem Cells International. 2011: 139851. doi:10.4061/2011/139851. PMC 3189604. PMID 22007239.
- ↑ Smith BW, Rozelle SS, Leung A, Ubellacker J, Parks A, Nah SK, et al. (July 2013). "The aryl hydrocarbon receptor directs hematopoietic progenitor cell expansion and differentiation". Blood. 122 (3): 376–85. doi:10.1182/blood-2012-11-466722. PMC 3716202. PMID 23723449.
- ↑ Shah S, Huang X, Cheng L (March 2014). "Concise review: stem cell-based approaches to red blood cell production for transfusion". Stem Cells Translational Medicine. 3 (3): 346–55. doi:10.5966/sctm.2013-0054. PMC 3952923. PMID 24361925.
- 1 2 Rousseau GF, Mazurier C, Douay L (2016). "Culturing red blood cells from stem cells: a solution to present and future challenges of transfusion medicine?". ISBT Science Series. 11: 111–117. doi:10.1111/voxs.12235. S2CID 88245602.
- ↑ Giani FC, Fiorini C, Wakabayashi A, Ludwig LS, Salem RM, Jobaliya CD, et al. (January 2016). "Targeted Application of Human Genetic Variation Can Improve Red Blood Cell Production from Stem Cells". Cell Stem Cell. 18 (1): 73–78. doi:10.1016/j.stem.2015.09.015. PMC 4707983. PMID 26607381.
- ↑ Fujita A, Uchida N, Haro-Mora JJ, Winkler T, Tisdale J (June 2016). "β-Globin-Expressing Definitive Erythroid Progenitor Cells Generated from Embryonic and Induced Pluripotent Stem Cell-Derived Sacs". Stem Cells. 34 (6): 1541–52. doi:10.1002/stem.2335. PMC 4892957. PMID 26866725.
- ↑ Figueiredo C, Goudeva L, Horn PA, Eiz-Vesper B, Blasczyk R, Seltsam A (August 2010). "Generation of HLA-deficient platelets from hematopoietic progenitor cells". Transfusion. 50 (8): 1690–701. doi:10.1111/j.1537-2995.2010.02644.x. PMID 20412529. S2CID 31313322.
- ↑ Suzuki D, Flahou C, Yoshikawa N, Stirblyte I, Hayashi Y, Sawaguchi A, et al. (January 2020). "iPSC-Derived Platelets Depleted of HLA Class I Are Inert to Anti-HLA Class I and Natural Killer Cell Immunity". Stem Cell Reports. Elsevier BV. 14 (1): 49–59. doi:10.1016/j.stemcr.2019.11.011. PMC 6962657. PMID 31883921.
- ↑ Nakamura S, Takayama N, Hirata S, Seo H, Endo H, Ochi K, et al. (April 2014). "Expandable megakaryocyte cell lines enable clinically applicable generation of platelets from human induced pluripotent stem cells". Cell Stem Cell. 14 (4): 535–48. doi:10.1016/j.stem.2014.01.011. PMID 24529595.
- ↑ Moreau T, Evans AL, Vasquez L, Tijssen MR, Yan Y, Trotter MW, et al. (April 2016). "Large-scale production of megakaryocytes from human pluripotent stem cells by chemically defined forward programming". Nature Communications. 7: 11208. Bibcode:2016NatCo...711208M. doi:10.1038/ncomms11208. hdl:1810/253670. PMC 4829662. PMID 27052461.
- ↑ Thon JN, Medvetz DA, Karlsson SM, Italiano JE (June 2015). "Road blocks in making platelets for transfusion". Journal of Thrombosis and Haemostasis. 13 (Suppl 1): S55-62. doi:10.1111/jth.12942. PMC 5565795. PMID 26149051.
- ↑ Riddell SR, Greenberg PD (1995). "Principles for adoptive T cell therapy of human viral diseases". Annual Review of Immunology. 13: 545–86. doi:10.1146/annurev.iy.13.040195.002553. PMID 7612234.
- 1 2 Nishimura T, Kaneko S, Kawana-Tachikawa A, Tajima Y, Goto H, Zhu D, et al. (January 2013). "Generation of rejuvenated antigen-specific T cells by reprogramming to pluripotency and redifferentiation". Cell Stem Cell. 12 (1): 114–26. doi:10.1016/j.stem.2012.11.002. PMID 23290140.
- ↑ Vizcardo R, Masuda K, Yamada D, Ikawa T, Shimizu K, Fujii S, et al. (January 2013). "Regeneration of human tumor antigen-specific T cells from iPSCs derived from mature CD8(+) T cells". Cell Stem Cell. 12 (1): 31–6. doi:10.1016/j.stem.2012.12.006. PMID 23290135.
- 1 2 Lei F, Haque R, Xiong X, Song J (May 2012). "Directed differentiation of induced pluripotent stem cells towards T lymphocytes". Journal of Visualized Experiments (63): e3986. doi:10.3791/3986. PMC 3389997. PMID 22617911.
- ↑ Sadelain M, Brentjens R, Rivière I (April 2013). "The basic principles of chimeric antigen receptor design". Cancer Discovery. 3 (4): 388–98. doi:10.1158/2159-8290.CD-12-0548. PMC 3667586. PMID 23550147.
- ↑ Themeli M, Kloss CC, Ciriello G, Fedorov VD, Perna F, Gonen M, Sadelain M (October 2013). "Generation of tumor-targeted human T lymphocytes from induced pluripotent stem cells for cancer therapy". Nature Biotechnology. 31 (10): 928–33. doi:10.1038/nbt.2678. PMC 5722218. PMID 23934177.
- ↑ Pilones KA, Aryankalayil J, Demaria S (2012). "Invariant NKT cells as novel targets for immunotherapy in solid tumors". Clinical & Developmental Immunology. 2012: 720803. doi:10.1155/2012/720803. PMC 3483734. PMID 23118781.
- ↑ Watarai H, Yamada D, Fujii S, Taniguchi M, Koseki H (June 2012). "Induced pluripotency as a potential path towards iNKT cell-mediated cancer immunotherapy". International Journal of Hematology. 95 (6): 624–31. doi:10.1007/s12185-012-1091-0. PMID 22592322. S2CID 20818388.
- ↑ Haruta M, Tomita Y, Yuno A, Matsumura K, Ikeda T, Takamatsu K, et al. (May 2013). "TAP-deficient human iPS cell-derived myeloid cell lines as unlimited cell source for dendritic cell-like antigen-presenting cells". Gene Therapy. 20 (5): 504–13. doi:10.1038/gt.2012.59. PMID 22875043.
- ↑ Xie H, Ye M, Feng R, Graf T (May 2004). "Stepwise reprogramming of B cells into macrophages". Cell. 117 (5): 663–76. doi:10.1016/S0092-8674(04)00419-2. PMID 15163413. S2CID 17156192.
Bussmann LH, Schubert A, Vu Manh TP, De Andres L, Desbordes SC, Parra M, et al. (November 2009). "A robust and highly efficient immune cell reprogramming system". Cell Stem Cell. 5 (5): 554–66. doi:10.1016/j.stem.2009.10.004. PMID 19896445. - ↑ Hanna J, Markoulaki S, Schorderet P, Carey BW, Beard C, Wernig M, et al. (April 2008). "Direct reprogramming of terminally differentiated mature B lymphocytes to pluripotency". Cell. 133 (2): 250–64. doi:10.1016/j.cell.2008.03.028. PMC 2615249. PMID 18423197.
- ↑ Di Stefano B, Sardina JL, van Oevelen C, Collombet S, Kallin EM, Vicent GP, et al. (February 2014). "C/EBPα poises B cells for rapid reprogramming into induced pluripotent stem cells". Nature. 506 (7487): 235–9. Bibcode:2014Natur.506..235D. doi:10.1038/nature12885. hdl:10803/283484. PMID 24336202. S2CID 4468460.
- ↑ Rapino F, Robles EF, Richter-Larrea JA, Kallin EM, Martinez-Climent JA, Graf T (April 2013). "C/EBPα induces highly efficient macrophage transdifferentiation of B lymphoma and leukemia cell lines and impairs their tumorigenicity". Cell Reports. 3 (4): 1153–63. doi:10.1016/j.celrep.2013.03.003. PMID 23545498.
- ↑ Guo J, Feng Y, Barnes P, Huang FF, Idell S, Su DM, Shams H (2012). "Deletion of FoxN1 in the thymic medullary epithelium reduces peripheral T cell responses to infection and mimics changes of aging". PLOS ONE. 7 (4): e34681. Bibcode:2012PLoSO...734681G. doi:10.1371/journal.pone.0034681. PMC 3326029. PMID 22514652.
- ↑ Sun L, Guo J, Brown R, Amagai T, Zhao Y, Su DM (June 2010). "Declining expression of a single epithelial cell-autonomous gene accelerates age-related thymic involution". Aging Cell. 9 (3): 347–57. doi:10.1111/j.1474-9726.2010.00559.x. PMC 2894280. PMID 20156205.
- ↑ Bredenkamp N, Nowell CS, Blackburn CC (April 2014). "Regeneration of the aged thymus by a single transcription factor". Development. 141 (8): 1627–37. doi:10.1242/dev.103614. PMC 3978836. PMID 24715454.
- ↑ Oh, J., Wang, W., Thomas, R., & Su, D. M. (2020). Thymic rejuvenation via induced thymic epithelial cells (iTECs) from FOXN1-overexpressing fibroblasts to counteract inflammaging. bioRxiv. https://doi.org/10.1101/2020.03.17.995357
- 1 2 Peng Y, Huang S, Cheng B, Nie X, Enhe J, Feng C, Fu X (January 2013). "Mesenchymal stem cells: a revolution in therapeutic strategies of age-related diseases". Ageing Research Reviews. 12 (1): 103–15. doi:10.1016/j.arr.2012.04.005. PMID 22569401. S2CID 44484625.
- 1 2 Bieback K, Kern S, Kocaömer A, Ferlik K, Bugert P (2008). "Comparing mesenchymal stromal cells from different human tissues: bone marrow, adipose tissue and umbilical cord blood". Bio-Medical Materials and Engineering. 18 (1 Suppl): S71-6. PMID 18334717.
- ↑ Efimenko A, Dzhoyashvili N, Kalinina N, Kochegura T, Akchurin R, Tkachuk V, Parfyonova Y (January 2014). "Adipose-derived mesenchymal stromal cells from aged patients with coronary artery disease keep mesenchymal stromal cell properties but exhibit characteristics of aging and have impaired angiogenic potential". Stem Cells Translational Medicine. 3 (1): 32–41. doi:10.5966/sctm.2013-0014. PMC 3902283. PMID 24353175.
- ↑ Stolzing A, Jones E, McGonagle D, Scutt A (March 2008). "Age-related changes in human bone marrow-derived mesenchymal stem cells: consequences for cell therapies". Mechanisms of Ageing and Development. 129 (3): 163–73. doi:10.1016/j.mad.2007.12.002. PMID 18241911. S2CID 25822581.
- ↑ Eberle I, Moslem M, Henschler R, Cantz T (2012). "Engineered MSCS from Patient-Specific iPS Cells". Mesenchymal Stem Cells - Basics and Clinical Application II. Advances in Biochemical Engineering/Biotechnology. Vol. 130. pp. 1–17. doi:10.1007/10_2012_156. ISBN 978-3-642-37943-7. PMID 22915200.
- ↑ Chen YS, Pelekanos RA, Ellis RL, Horne R, Wolvetang EJ, Fisk NM (February 2012). "Small molecule mesengenic induction of human induced pluripotent stem cells to generate mesenchymal stem/stromal cells". Stem Cells Translational Medicine. 1 (2): 83–95. doi:10.5966/sctm.2011-0022. PMC 3659681. PMID 23197756.
- ↑ Hynes K, Menicanin D, Han J, Marino V, Mrozik K, Gronthos S, Bartold PM (September 2013). "Mesenchymal stem cells from iPS cells facilitate periodontal regeneration". Journal of Dental Research. 92 (9): 833–9. doi:10.1177/0022034513498258. PMID 23884555. S2CID 20811301.
- ↑ Hynes K, Gronthos S, Bartold PM (March 1, 2014). "iPSC for Dental Tissue Regeneration". Current Oral Health Reports. 1 (1): 9–15. doi:10.1007/s40496-013-0001-8.
- ↑ Lai PL, Lin H, Chen SF, Yang SC, Hung KH, Chang CF, et al. (March 2017). "Efficient Generation of Chemically Induced Mesenchymal Stem Cells from Human Dermal Fibroblasts". Scientific Reports. 7: 44534. Bibcode:2017NatSR...744534L. doi:10.1038/srep44534. PMC 5356011. PMID 28303927.
- ↑ Lai RC, Yeo RW, Tan SS, Zhang B, Yin Y, Sze NS, Choo A, Lim SK (2013). "Mesenchymal Stem Cell Exosomes: The Future MSC-Based Therapy?". Mesenchymal Stem Cell Therapy. pp. 39–61. doi:10.1007/978-1-62703-200-1_3. ISBN 978-1-62703-199-8.
Lai RC, Yeo RW, Tan KH, Lim SK (2013). "Exosomes for drug delivery - a novel application for the mesenchymal stem cell". Biotechnology Advances. 31 (5): 543–51. doi:10.1016/j.biotechadv.2012.08.008. PMID 22959595.
Kosaka N, Takeshita F, Yoshioka Y, Hagiwara K, Katsuda T, Ono M, Ochiya T (March 2013). "Exosomal tumor-suppressive microRNAs as novel cancer therapy: "exocure" is another choice for cancer treatment". Advanced Drug Delivery Reviews. 65 (3): 376–82. doi:10.1016/j.addr.2012.07.011. PMID 22841506. - ↑ Jumabay M, Abdmaulen R, Ly A, Cubberly MR, Shahmirian LJ, Heydarkhan-Hagvall S, et al. (February 2014). "Pluripotent stem cells derived from mouse and human white mature adipocytes". Stem Cells Translational Medicine. 3 (2): 161–71. doi:10.5966/sctm.2013-0107. PMC 3925054. PMID 24396033.
- ↑ Poloni A, Maurizi G, Leoni P, Serrani F, Mancini S, Frontini A, et al. (May 2012). "Human dedifferentiated adipocytes show similar properties to bone marrow-derived mesenchymal stem cells". Stem Cells. 30 (5): 965–74. doi:10.1002/stem.1067. hdl:11566/65996. PMID 22367678. S2CID 25979840.
- ↑ Shen JF, Sugawara A, Yamashita J, Ogura H, Sato S (July 2011). "Dedifferentiated fat cells: an alternative source of adult multipotent cells from the adipose tissues". International Journal of Oral Science. 3 (3): 117–24. doi:10.4248/IJOS11044. PMC 3470092. PMID 21789960.
- ↑ Melief SM, Zwaginga JJ, Fibbe WE, Roelofs H (June 2013). "Adipose tissue-derived multipotent stromal cells have a higher immunomodulatory capacity than their bone marrow-derived counterparts". Stem Cells Translational Medicine. 2 (6): 455–63. doi:10.5966/sctm.2012-0184. PMC 3673757. PMID 23694810.
- ↑ Cheng A, Hardingham TE, Kimber SJ (August 2014). "Generating cartilage repair from pluripotent stem cells". Tissue Engineering. Part B, Reviews. 20 (4): 257–66. doi:10.1089/ten.teb.2012.0757. PMC 4123466. PMID 23957872.
- ↑ Outani H, Okada M, Yamashita A, Nakagawa K, Yoshikawa H, Tsumaki N (2013). "Direct induction of chondrogenic cells from human dermal fibroblast culture by defined factors". PLOS ONE. 8 (10): e77365. Bibcode:2013PLoSO...877365O. doi:10.1371/journal.pone.0077365. PMC 3797820. PMID 24146984.
- 1 2 Crompton JG, Rao M, Restifo NP (January 2013). "Memoirs of a reincarnated T cell". Cell Stem Cell. 12 (1): 6–8. doi:10.1016/j.stem.2012.12.009. PMC 6352969. PMID 23290132.
- ↑ Tan HK, Toh CX, Ma D, Yang B, Liu TM, Lu J, et al. (May 2014). "Human finger-prick induced pluripotent stem cells facilitate the development of stem cell banking". Stem Cells Translational Medicine. 3 (5): 586–98. doi:10.5966/sctm.2013-0195. PMC 4006490. PMID 24646489.
- ↑ Okita K, Yamakawa T, Matsumura Y, Sato Y, Amano N, Watanabe A, et al. (March 2013). "An efficient nonviral method to generate integration-free human-induced pluripotent stem cells from cord blood and peripheral blood cells". Stem Cells. 31 (3): 458–66. doi:10.1002/stem.1293. PMID 23193063. S2CID 206512330.
- ↑ Geti I, Ormiston ML, Rouhani F, Toshner M, Movassagh M, Nichols J, et al. (December 2012). "A practical and efficient cellular substrate for the generation of induced pluripotent stem cells from adults: blood-derived endothelial progenitor cells". Stem Cells Translational Medicine. 1 (12): 855–65. doi:10.5966/sctm.2012-0093. PMC 3659672. PMID 23283547.
- ↑ Staerk J, Dawlaty MM, Gao Q, Maetzel D, Hanna J, Sommer CA, et al. (July 2010). "Reprogramming of human peripheral blood cells to induced pluripotent stem cells". Cell Stem Cell. 7 (1): 20–4. doi:10.1016/j.stem.2010.06.002. PMC 2917234. PMID 20621045.
- ↑ Park TS, Huo JS, Peters A, Talbot CC, Verma K, Zimmerlin L, et al. (2012). "Growth factor-activated stem cell circuits and stromal signals cooperatively accelerate non-integrated iPSC reprogramming of human myeloid progenitors". PLOS ONE. 7 (8): e42838. Bibcode:2012PLoSO...742838P. doi:10.1371/journal.pone.0042838. PMC 3414503. PMID 22905176.
- ↑ Yoshikawa K, Naitoh M, Kubota H, Ishiko T, Aya R, Yamawaki S, Suzuki S (February 2013). "Multipotent stem cells are effectively collected from adult human cheek skin". Biochemical and Biophysical Research Communications. 431 (1): 104–10. doi:10.1016/j.bbrc.2012.12.069. hdl:2433/174817. PMID 23268344.
- ↑ Zhou T, Benda C, Duzinger S, Huang Y, Li X, Li Y, et al. (July 2011). "Generation of induced pluripotent stem cells from urine". Journal of the American Society of Nephrology. 22 (7): 1221–8. doi:10.1681/ASN.2011010106. PMC 3137570. PMID 21636641.
- ↑ Zhou T, Benda C, Dunzinger S, Huang Y, Ho JC, Yang J, et al. (December 2012). "Generation of human induced pluripotent stem cells from urine samples". Nature Protocols. 7 (12): 2080–9. doi:10.1038/nprot.2012.115. PMID 23138349. S2CID 205465442.
- ↑ Wang L, Wang L, Huang W, Su H, Xue Y, Su Z, et al. (January 2013). "Generation of integration-free neural progenitor cells from cells in human urine". Nature Methods. 10 (1): 84–9. doi:10.1038/nmeth.2283. hdl:10397/32665. PMID 23223155. S2CID 1475859.
- ↑ Cai J, Zhang Y, Liu P, Chen S, Wu X, Sun Y, et al. (2013). "Generation of tooth-like structures from integration-free human urine induced pluripotent stem cells". Cell Regeneration. 2 (1): 6. doi:10.1186/2045-9769-2-6. PMC 4230506. PMID 25408878.
- ↑ Bharadwaj S, Liu G, Shi Y, Wu R, Yang B, He T, et al. (September 2013). "Multipotential differentiation of human urine-derived stem cells: potential for therapeutic applications in urology". Stem Cells. 31 (9): 1840–56. doi:10.1002/stem.1424. PMID 23666768. S2CID 2380066.
- ↑ Wang Y, Liu J, Tan X, Li G, Gao Y, Liu X, et al. (August 2013). "Induced pluripotent stem cells from human hair follicle mesenchymal stem cells". Stem Cell Reviews and Reports. 9 (4): 451–60. doi:10.1007/s12015-012-9420-5. PMC 3742959. PMID 23242965.
- ↑ Schnabel LV, Abratte CM, Schimenti JC, Southard TL, Fortier LA (August 2012). "Genetic background affects induced pluripotent stem cell generation". Stem Cell Research & Therapy. 3 (4): 30. doi:10.1186/scrt121. PMC 3580468. PMID 22862934.
- ↑ Panopoulos AD, Ruiz S, Yi F, Herrerías A, Batchelder EM, Izpisua Belmonte JC (2011). "Rapid and highly efficient generation of induced pluripotent stem cells from human umbilical vein endothelial cells". PLOS ONE. 6 (5): e19743. Bibcode:2011PLoSO...619743P. doi:10.1371/journal.pone.0019743. PMC 3095638. PMID 21603572.
- 1 2 Polo JM, Liu S, Figueroa ME, Kulalert W, Eminli S, Tan KY, et al. (August 2010). "Cell type of origin influences the molecular and functional properties of mouse induced pluripotent stem cells". Nature Biotechnology. 28 (8): 848–55. doi:10.1038/nbt.1667. PMC 3148605. PMID 20644536.
- ↑ Miura K, Okada Y, Aoi T, Okada A, Takahashi K, Okita K, et al. (August 2009). "Variation in the safety of induced pluripotent stem cell lines". Nature Biotechnology. 27 (8): 743–5. doi:10.1038/nbt.1554. hdl:2433/120546. PMID 19590502. S2CID 1372523.
- ↑ Liang Y, Zhang H, Feng QS, Cai MB, Deng W, Qin D, et al. (April 2013). "The propensity for tumorigenesis in human induced pluripotent stem cells is related with genomic instability". Chinese Journal of Cancer. 32 (4): 205–12. doi:10.5732/cjc.012.10065. PMC 3845575. PMID 22704487.
- ↑ Kim K, Doi A, Wen B, Ng K, Zhao R, Cahan P, et al. (September 2010). "Epigenetic memory in induced pluripotent stem cells". Nature. 467 (7313): 285–90. Bibcode:2010Natur.467..285K. doi:10.1038/nature09342. PMC 3150836. PMID 20644535.
- ↑ Kim K, Zhao R, Doi A, Ng K, Unternaehrer J, Cahan P, et al. (November 2011). "Donor cell type can influence the epigenome and differentiation potential of human induced pluripotent stem cells". Nature Biotechnology. 29 (12): 1117–9. doi:10.1038/nbt.2052. PMC 3357310. PMID 22119740.
- ↑ Bar-Nur O, Russ HA, Efrat S, Benvenisty N (July 2011). "Epigenetic memory and preferential lineage-specific differentiation in induced pluripotent stem cells derived from human pancreatic islet beta cells". Cell Stem Cell. 9 (1): 17–23. doi:10.1016/j.stem.2011.06.007. PMID 21726830.
- ↑ Denker HW (December 2012). "Time to reconsider stem cell induction strategies". Cells. 1 (4): 1293–312. doi:10.3390/cells1041293. PMC 3901125. PMID 24710555.
Further reading
- Scudellari M (June 2016). "How iPS cells changed the world". Nature. 534 (7607): 310–2. Bibcode:2016Natur.534..310S. doi:10.1038/534310a. PMID 27306170.
- Turksen K, Nagy A (2016). Induced Pluripotent Stem (iPS) Cells. Methods in Molecular Biology. Vol. 1357. doi:10.1007/978-1-4939-3055-5. ISBN 978-1-4939-3054-8. S2CID 46010857.
- Li M, Izpisua Belmonte JC (September 2016). "Looking to the future following 10 years of induced pluripotent stem cell technologies". Nature Protocols. 11 (9): 1579–85. doi:10.1016/j.cell.2016.08.055. PMID 27490631.
- Srivastava D, DeWitt N (September 2016). "In Vivo Cellular Reprogramming: The Next Generation". Cell. 166 (6): 1386–1396. doi:10.1016/j.cell.2016.08.055. PMC 6234007. PMID 27610565.
- Tabar V, Studer L (February 2014). "Pluripotent stem cells in regenerative medicine: challenges and recent progress". Nature Reviews. Genetics. 15 (2): 82–92. doi:10.1038/nrg3563. PMC 4539940. PMID 24434846.
- Tan Y, Ooi S, Wang L (January 2014). "Immunogenicity and tumorigenicity of pluripotent stem cells and their derivatives: genetic and epigenetic perspectives". Current Stem Cell Research & Therapy. 9 (1): 63–72. doi:10.2174/1574888x113086660068. PMC 3873036. PMID 24160683.
- Yamanaka S (June 2012). "Induced pluripotent stem cells: past, present, and future". Cell Stem Cell. 10 (6): 678–684. doi:10.1016/j.stem.2012.05.005. PMID 22704507.
- Takahashi K, Yamanaka S (June 2013). "Induced pluripotent stem cells in medicine and biology". Development. 140 (12): 2457–61. doi:10.1242/dev.092551. PMID 23715538.
- Asuelime GE, Shi Y (August 2012). "A case of cellular alchemy: lineage reprogramming and its potential in regenerative medicine". Journal of Molecular Cell Biology. 4 (4): 190–6. doi:10.1093/jmcb/mjs005. PMC 3408064. PMID 22371436.
- Lensch MW, Mummery CL (June 2013). "From stealing fire to cellular reprogramming: a scientific history leading to the 2012 Nobel Prize". Stem Cell Reports. 1 (1): 5–17. doi:10.1016/j.stemcr.2013.05.001. PMC 3757737. PMID 24052937.
- Zhou Q (October 2013). "Induced pluripotent stem cells: current progress and future perspectives. Preface". Genomics, Proteomics & Bioinformatics. 11 (5): 257–8. doi:10.1016/j.gpb.2013.10.001. PMC 4357784. PMID 24121127.
- Lin J, Li MR, Ti DD, Chen MX, Hao HJ, Zhao YL, et al. (January 2013). "Microenvironment-evoked cell lineage conversion: Shifting the focus from internal reprogramming to external forcing". Ageing Research Reviews. 12 (1): 29–38. doi:10.1016/j.arr.2012.04.002. PMID 22561469. S2CID 40367513.
- Takahashi K (February 2014). "Cellular reprogramming". Cold Spring Harbor Perspectives in Biology. 6 (2): a018606. doi:10.1101/cshperspect.a018606. PMC 3941237. PMID 24492711.
- Nobel Prize in Physiology or Medicine 2012 Awarded for Discovery That Mature Cells Can Be Reprogrammed to Become Pluripotent
- Hussein SM, Nagy AA (October 2012). "Progress made in the reprogramming field: new factors, new strategies and a new outlook". Current Opinion in Genetics & Development. 22 (5): 435–43. doi:10.1016/j.gde.2012.08.007. PMID 22959308.
- Zhang Y, Yao L, Yu X, Ou J, Hui N, Liu S (December 2012). "A poor imitation of a natural process: a call to reconsider the iPSC engineering technique". Cell Cycle. 11 (24): 4536–44. doi:10.4161/cc.22575. PMC 3562298. PMID 23114619.
- Luni C, Giulitti S, Serena E, Ferrari L, Zambon A, Gagliano O, et al. (May 2016). "High-efficiency cellular reprogramming with microfluidics". Nature Methods. 13 (5): 446–52. doi:10.1038/nmeth.3832. PMID 27088312. S2CID 5462386. a 50-fold increase in efficiency; small volumes; differentiation into desired cells in the same platform
- Mochiduki Y, Okita K (June 2012). "Methods for iPS cell generation for basic research and clinical applications". Biotechnology Journal. 7 (6): 789–97. doi:10.1002/biot.201100356. PMID 22378737. S2CID 22317543.
- Madonna R (October 2012). "Human-induced pluripotent stem cells: in quest of clinical applications". Molecular Biotechnology. 52 (2): 193–203. doi:10.1007/s12033-012-9504-0. PMID 22302314. S2CID 9920142.
- Lorenzo IM, Fleischer A, Bachiller D (August 2013). "Generation of mouse and human induced pluripotent stem cells (iPSC) from primary somatic cells". Stem Cell Reviews and Reports. 9 (4): 435–50. doi:10.1007/s12015-012-9412-5. hdl:10261/98876. PMID 23104133. S2CID 2225015.
- "Stem Cell Reprogramming Tools". Invitrogen.
Detailed protocols for reprogramming and for analysis of iPSCs
- Buganim Y, Faddah DA, Jaenisch R (June 2013). "Mechanisms and models of somatic cell reprogramming". Nature Reviews. Genetics. 14 (6): 427–39. doi:10.1038/nrg3473. PMC 4060150. PMID 23681063.
| Sources/types | |
|---|---|
| Cell potency |
|
| Related articles | |
| |