| Betacoronavirus NS8 protein | |||||||||
|---|---|---|---|---|---|---|---|---|---|
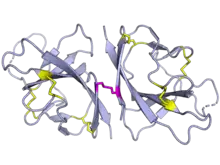 The SARS-CoV-2 ORF8 protein dimer. Internal disulfide bonds are shown in yellow and the intermolecular disulfide is shown in magenta. From PDB: 7JTL.[1] | |||||||||
| Identifiers | |||||||||
| Symbol | bCoV_NS8 | ||||||||
| Pfam | PF12093 | ||||||||
| InterPro | IPR022722 | ||||||||
| |||||||||
ORF8 is a gene that encodes a viral accessory protein, Betacoronavirus NS8 protein, in coronaviruses of the subgenus Sarbecovirus. It is one of the least well conserved and most variable parts of the genome.[2][3][4][5] In some viruses, a deletion splits the region into two smaller open reading frames, called ORF8a and ORF8b - a feature present in many SARS-CoV viral isolates from later in the SARS epidemic, as well as in some bat coronaviruses.[4][3] For this reason the full-length gene and its protein are sometimes called ORF8ab.[3][6] The full-length gene, exemplified in SARS-CoV-2, encodes a protein with an immunoglobulin domain of unknown function, possibly involving interactions with the host immune system.[4][3][1] It is similar in structure to the ORF7a protein, suggesting it may have originated through gene duplication.[7][8]
Structure
ORF8 in SARS-CoV-2 encodes a protein of 121 amino acid residues with an N-terminal signal sequence.[4] ORF8 forms a dimer that is covalently linked by disulfide bonds.[1] It has an immunoglobulin-like domain with distant similarity to the ORF7a protein.[1][2] Despite a similar overall fold, an insertion in ORF8 likely is responsible for different protein-protein interactions and creates an additional dimerization interface.[1][2] Unlike ORF7a, ORF8 lacks a transmembrane helix and is therefore not a transmembrane protein,[1][4] though it has been suggested it might have a membrane-anchored form.[3]
ORF8 in SARS-CoV and SARS-CoV-2 are very divergent, with less than 20% sequence identity.[1] The full-length ORF8 in SARS-CoV encodes a protein of 122 residues. In many SARS-CoV isolates it is split into ORF8a and ORF8b, separately expressing 39-residue ORF8a and 84-residue ORF8b proteins.[6] It has been suggested that the ORF8a and ORF8b proteins may form a protein complex.[2][9] The cysteine residue responsible for dimerization of the SARS-CoV-2 protein is not conserved in the SARS-CoV sequence.[1] The ORF8ab protein has also been reported to form disulfide-linked multimers.[10]
Post-translational modifications
The full-length SARS-CoV ORF8ab protein is post-translationally modified by N-glycosylation,[6] which is predicted to be conserved in the SARS-CoV-2 protein.[1] Under experimental conditions, both 8b and 8ab are ubiquitinated.[6]
Expression and localization
Along with the genes for other accessory proteins, the ORF8 gene is located near those encoding the structural proteins, at the 5' end of the coronavirus RNA genome. Along with ORF6, ORF7a, and ORF7b, ORF8 is located between the membrane (M) and nucleocapsid (N) genes.[6][4] The SARS-CoV-2 ORF8 protein has a signal sequence for trafficking to the endoplasmic reticulum (ER)[4] and has been experimentally localized to the ER.[11] It is probably a secreted protein.[4][3]
There are variable reports in the literature regarding the localization of SARS-CoV ORF8a, ORF8b, or ORF8ab proteins.[6] It is unclear if ORF8b is expressed at significant levels under natural conditions.[10][12] The full-length ORF8ab appears to localize to the ER.[12]
Function
The function of the ORF8 protein is unknown. It is not essential for viral replication in either SARS-CoV[6] or SARS-CoV-2,[4] though there is conflicting evidence on whether loss of ORF8 affects the efficiency of viral replication.[13]
A function often suggested for ORF8 protein is interacting with the host immune system.[13] The SARS-CoV-2 protein is thought to have a role in immunomodulation via immune evasion or suppressing host immune responses.[4][1][3] It has been reported to be a type I interferon antagonist and to downregulate class I MHC.[4][3] The SARS-CoV-2 ORF8 protein is highly immunogenic and high levels of antibodies to the protein have been found in patients with or recovered from COVID-19.[4][14] A study indicates that ORF8 is a transcription inhibitor.[15][16]
It has been suggested that the SARS-CoV ORF8a protein assembles into multimers and forms a viroporin.[17]
Evolution

The evolutionary history of ORF8 is complex. It is among the least conserved regions of the Sarbecovirus genome.[3][2][4] It is subject to frequent mutations and deletions, and has been described as "hypervariable" and a recombination hotspot.[3] It has been suggested that RNA secondary structures in the region are associated with genomic instability.[3][19]
In SARS-CoV, the ORF8 region is thought to have originated through recombination among ancestral bat coronaviruses.[3][6][5][20] Among the most distinctive features of this region in SARS-CoV is the emergence of a 29-nucleotide deletion that split the full-length open reading frame into two smaller ORFs, ORF8a and ORF8b. Viral isolates from early in the SARS epidemic have a full-length, intact ORF8, but the split structure emerged later in the epidemic.[3][6] Similar split structures have since been observed in bat coronaviruses.[21] Mutations and deletions have also been seen in SARS-CoV-2 variants.[2][19] Based on observations in SARS-CoV, it has been suggested that changes in ORF8 may be related to host adaptation, but it is possible that ORF8 does not affect fitness in human hosts.[19][5] In SARS-CoV, a high dN/dS ratio has been observed in ORF8, consistent with positive selection or with relaxed selection.[5]
ORF8 encodes a protein whose immunoglobulin domain (Ig) has distant similarity to that of ORF7a.[1] It has been suggested that ORF8 likely have evolved from ORF7a through gene duplication,[2][7][8] though some bioinformatics analyses suggest the similarity may be too low to support duplication, which is relatively uncommon in viruses.[19] Immunoglobulin domains are uncommon in coronaviruses; other than the subset of betacoronaviruses with ORF8 and ORF7a, only a small number of bat alphacoronaviruses have been identified as containing likely Ig domains, while they are absent from gammacoronaviruses and deltacoronaviruses.[2][8] ORF8 is notably absent in MERS-CoV.[8] The beta and alpha Ig domains may be independent acquisitions, where ORF8 and ORF7a may have been acquired from host proteins.[2] It is also possible that the absence of ORF8 reflects gene loss in those lineages.[8]
References
- 1 2 3 4 5 6 7 8 9 10 11 12 Flower TG, Buffalo CZ, Hooy RM, Allaire M, Ren X, Hurley JH (January 2021). "Structure of SARS-CoV-2 ORF8, a rapidly evolving immune evasion protein". Proceedings of the National Academy of Sciences of the United States of America. 118 (2): e2021785118. Bibcode:2021PNAS..11821785F. doi:10.1073/pnas.2021785118. PMC 7812859. PMID 33361333.
- 1 2 3 4 5 6 7 8 9 Tan Y, Schneider T, Leong M, Aravind L, Zhang D (May 2020). "Novel Immunoglobulin Domain Proteins Provide Insights into Evolution and Pathogenesis of SARS-CoV-2-Related Viruses". mBio. 11 (3). doi:10.1128/mBio.00760-20. PMC 7267882. PMID 32471829.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 Zinzula L (January 2021). "Lost in deletion: The enigmatic ORF8 protein of SARS-CoV-2". Biochemical and Biophysical Research Communications. 538: 116–124. doi:10.1016/j.bbrc.2020.10.045. PMC 7577707. PMID 33685621.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 Redondo N, Zaldívar-López S, Garrido JJ, Montoya M (7 July 2021). "SARS-CoV-2 Accessory Proteins in Viral Pathogenesis: Knowns and Unknowns". Frontiers in Immunology. 12: 708264. doi:10.3389/fimmu.2021.708264. hdl:10261/249329. PMC 8293742. PMID 34305949.
- 1 2 3 4 Forni D, Cagliani R, Clerici M, Sironi M (January 2017). "Molecular Evolution of Human Coronavirus Genomes". Trends in Microbiology. 25 (1): 35–48. doi:10.1016/j.tim.2016.09.001. PMC 7111218. PMID 27743750.
- 1 2 3 4 5 6 7 8 9 Liu DX, Fung TS, Chong KK, Shukla A, Hilgenfeld R (September 2014). "Accessory proteins of SARS-CoV and other coronaviruses". Antiviral Research. 109: 97–109. doi:10.1016/j.antiviral.2014.06.013. PMC 7113789. PMID 24995382.
- 1 2 Mariano G, Farthing RJ, Lale-Farjat SL, Bergeron JR (17 December 2020). "Structural Characterization of SARS-CoV-2: Where We Are, and Where We Need to Be". Frontiers in Molecular Biosciences. 7: 605236. doi:10.3389/fmolb.2020.605236. PMC 7773825. PMID 33392262.
- 1 2 3 4 5 Neches RY, Kyrpides NC, Ouzounis CA (January 2021). "Atypical Divergence of SARS-CoV-2 Orf8 from Orf7a within the Coronavirus Lineage Suggests Potential Stealthy Viral Strategies in Immune Evasion". mBio. 12 (1). doi:10.1128/mBio.03014-20. PMC 7845636. PMID 33468697.
- ↑ von Brunn A, Teepe C, Simpson JC, Pepperkok R, Friedel CC, Zimmer R, et al. (May 2007). "Analysis of intraviral protein-protein interactions of the SARS coronavirus ORFeome". PLOS ONE. 2 (5): e459. Bibcode:2007PLoSO...2..459V. doi:10.1371/journal.pone.0000459. PMC 1868897. PMID 17520018.
- 1 2 Oostra M, de Haan CA, Rottier PJ (December 2007). "The 29-nucleotide deletion present in human but not in animal severe acute respiratory syndrome coronaviruses disrupts the functional expression of open reading frame 8". Journal of Virology. 81 (24): 13876–13888. doi:10.1128/JVI.01631-07. PMC 2168875. PMID 17928347.
- ↑ Zhang J, Cruz-Cosme R, Zhuang MW, Liu D, Liu Y, Teng S, et al. (November 2020). "A systemic and molecular study of subcellular localization of SARS-CoV-2 proteins". Signal Transduction and Targeted Therapy. 5 (1): 269. doi:10.1038/s41392-020-00372-8. PMC 7670843. PMID 33203855.
- 1 2 McBride R, Fielding BC (November 2012). "The role of severe acute respiratory syndrome (SARS)-coronavirus accessory proteins in virus pathogenesis". Viruses. 4 (11): 2902–2923. doi:10.3390/v4112902. PMC 3509677. PMID 23202509.
- 1 2 Mohammad S, Bouchama A, Mohammad Alharbi B, Rashid M, Saleem Khatlani T, Gaber NS, Malik SS (August 2020). "SARS-CoV-2 ORF8 and SARS-CoV ORF8ab: Genomic Divergence and Functional Convergence". Pathogens. 9 (9): 677. doi:10.3390/pathogens9090677. PMC 7558349. PMID 32825438.
- ↑ Hachim A, Kavian N, Cohen CA, Chin AW, Chu DK, Mok CK, et al. (October 2020). "ORF8 and ORF3b antibodies are accurate serological markers of early and late SARS-CoV-2 infection". Nature Immunology. 21 (10): 1293–1301. doi:10.1038/s41590-020-0773-7. PMID 32807944. S2CID 221136730.
- ↑ Kee J, Thudium S, Renner DM, Glastad K, Palozola K, Zhang Z, et al. (October 2022). "SARS-CoV-2 disrupts host epigenetic regulation via histone mimicry". Nature. 610 (7931): 381–388. Bibcode:2022Natur.610..381K. doi:10.1038/s41586-022-05282-z. PMC 9533993. PMID 36198800.
- ↑ Thomann L, Thiel V (October 2022). "SARS-CoV-2 mimics a host protein to bypass defences". Nature. 610 (7931): 262–263. Bibcode:2022Natur.610..262T. doi:10.1038/d41586-022-02930-2. PMID 36198813. S2CID 252737550.
- ↑ Chen CC, Krüger J, Sramala I, Hsu HJ, Henklein P, Chen YM, Fischer WB (February 2011). "ORF8a of SARS-CoV forms an ion channel: experiments and molecular dynamics simulations". Biochimica et Biophysica Acta (BBA) - Biomembranes. 1808 (2): 572–579. doi:10.1016/j.bbamem.2010.08.004. PMC 7094593. PMID 20708597.
- ↑ Zhou Z, Huang C, Zhou Z, Huang Z, Su L, Kang S, et al. (March 2021). "Structural insight reveals SARS-CoV-2 ORF7a as an immunomodulating factor for human CD14+ monocytes". iScience. 24 (3): 102187. Bibcode:2021iSci...24j2187Z. doi:10.1016/j.isci.2021.102187. PMC 7879101. PMID 33615195.
- 1 2 3 4 Pereira F (November 2020). "Evolutionary dynamics of the SARS-CoV-2 ORF8 accessory gene". Infection, Genetics and Evolution. 85: 104525. doi:10.1016/j.meegid.2020.104525. PMC 7467077. PMID 32890763.
- ↑ Lau SK, Feng Y, Chen H, Luk HK, Yang WH, Li KS, et al. (October 2015). "Severe Acute Respiratory Syndrome (SARS) Coronavirus ORF8 Protein Is Acquired from SARS-Related Coronavirus from Greater Horseshoe Bats through Recombination". Journal of Virology. 89 (20): 10532–10547. doi:10.1128/JVI.01048-15. PMC 4580176. PMID 26269185.
- ↑ Hu B, Zeng LP, Yang XL, Ge XY, Zhang W, Li B, et al. (November 2017). "Discovery of a rich gene pool of bat SARS-related coronaviruses provides new insights into the origin of SARS coronavirus". PLOS Pathogens. 13 (11): e1006698. doi:10.1371/journal.ppat.1006698. PMC 5708621. PMID 29190287.
| Viral structural protein |
|  |
|---|---|---|
| Viral nonstructural protein (expressed from ORF1ab) |
| |
| Viral accessory protein | ||
| RNA | ||