| Palaeoloxodon Temporal range: Middle | |
|---|---|
.JPG.webp) | |
| Skeleton of the straight-tusked elephant (Palaeoloxodon antiquus) at the paleontological museum of the Sapienza University of Rome | |
| Scientific classification | |
| Domain: | Eukaryota |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Mammalia |
| Order: | Proboscidea |
| Clade: | Elephantida |
| Superfamily: | Elephantoidea |
| Family: | Elephantidae |
| Genus: | †Palaeoloxodon Matsumoto, 1924[1] |
| Type species | |
| Elephas namadicus naumanni Makiyama, 1924 | |
| Species | |
|
See text | |
Palaeoloxodon is an extinct genus of elephant. The genus originated in Africa during the Early Pleistocene, and expanded into Eurasia at the beginning of the Middle Pleistocene. The genus contains some of the largest known species of elephants, over 4 metres (13 ft) tall at the shoulders, including the African Palaeoloxodon recki, the European straight-tusked elephant (Palaeoloxodon antiquus) and the South Asian Palaeoloxodon namadicus. P. namadicus has been suggested to be the largest known land mammal by some authors based on extrapolation from fragmentary remains, though these estimates are highly speculative.[2] In contrast, the genus also contains many species of dwarf elephants that evolved via insular dwarfism on islands in the Mediterranean, some only 1 metre (3.3 ft) in height, making them the smallest elephants known. The genus has a long and complex taxonomic history, and at various times, it has been considered to belong to Loxodonta or Elephas, but today is usually considered a valid and separate genus in its own right.
Taxonomy
In 1924, Matsumoto Hikoshichirō circumscribed Palaeoloxodon as a subgenus of Loxodonta (African elephants). It included the "E. antiquus—namadicus group", and he designated the Japanese "E. namadicus naumanni Mak." as its type species.[1]

Palaeoloxodon was later thought to be a subgenus or a synonym of Elephas (which includes the Asian elephant), but this was subsequently abandoned.[3] In 2016, a study of the straight-tusked elephant (P. antiquus) mitochondrial genome and part of the nuclear genome found that the mitochondrial sequences were nested within the diversity of those of the African forest elephant, Loxodonta cyclotis, with the partial nuclear genome supporting P. antiquus as more closely related to L. cyclotis than the African bush elephant, L. africana.[4] A later study published in 2018 by the same authors based on the complete nuclear genome revised these results, and suggested P. antiquus had a complex hybridization history, with around 60% of its nuclear genome coming from a lineage most closely related to modern African elephants but which diverged before the split between the two living species, around 5% from Mammuthus and 36% deriving from L. cyclotis. The ancestry from L. cyclotis was more closely related to modern West African populations of the forest elephant than to other forest elephant populations, while the mammoth ancestry was basal to the split between woolly and Columbian mammoths, probably from shortly after the split between the ancestors of mammoths and Asian elephants. The hybridisation probably took place in Africa, where Palaeoloxodon was dominant for most of the Pliocene and Early Pleistocene, with the mammoth hybridisation suggested to have taken place earlier than the hybridisation with forest elephants.[5]
Analysis of mitochondrial genomes, including Palaeoloxodon individuals from Northern China indicates Palaeoloxodon individuals harboured multiple separate mitochondrial genome lineages derived from African forest elephants, some being more closely related to some West African forest elephant groups than to others. It is unclear as to whether this is the result of multiple hybridisation events, or whether multiple mitochondrial lineages were introgressed in a single event. It has been found that mitochondrial genome of Chinese Palaeoloxodon specimens clustered with a P. antiquus individual from western Europe, which belonged to a separate clade than other sampled European P. antiquus specimens. The relatively low divergence between the mitochondrial genomes of the European P. antiquus individual and the Chinese Palaeoloxodon specimens may indicate that the populations of Palaeoloxodon across Eurasia maintained gene flow with each other, but this is uncertain.[6]
Diagram of the relationships of elephant mitochondrial genomes, after Lin et al. 2023:[6]
| |||||||||||||||||||||||||||||||||||||||||||||||||
Mainland species
- P. recki (Synonym:Elephas recki) (East Africa), the oldest species and ancestor of all later species
- P. jolensis (Synonym: Elephas iolensis) the last (late Middle-Late Pleistocene) representative of Palaeoloxodon in Africa
- P. antiquus (Synonym:Elephas antiquus) (Straight tusked elephant) (Europe, Western Asia)
- P. huaihoensis (China)
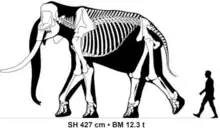
- P. namadicus (Synonym:Elephas namadicus)[7] (Indian subcontinent, possibly also elsewhere in Asia), the largest in its genus, and possibly the largest terrestrial mammal ever
- P. naumanni (Synonym:E. namadicus naumanni) (Japan, possibly also China and Korea),[8]
- ?P. turkmenicus known from a single specimen found in the Middle Pleistocene of Turkmenistan, with possibly attributable remains known from Kashmir, validity uncertain.[9]
Mediterranean island dwarfs
These Mediterranean insular dwarf elephant species are almost certainly descended from P. antiquus
- P. creutzburgi (Crete)
- P. xylophagou (Cyprus)
- P. cypriotes (Cyprus)
- P. lomolinoi (Naxos)
- P. tiliensis (Tilos)
- P. mnaidriensis (Sicily and Malta)
- P. falconeri (Sicily and Malta)
Description
Most species of Palaeoloxodon are noted for their distinctive parieto-occipital crests present at the top of the cranium, which was used to anchor the splenius as well as possibly the rhomboid muscles to support the skull, which is proportionally large in comparison to other elephants. The development of the crest is variable depending on the species, growth stage and gender, with females and juveniles having less developed or absent crests.[9] The skull is proportionally short and tall,[10] with the premaxillary bones containing the tusks being flared outwards. The tusks have relatively little curvature, and are proportionally large,[9] and somewhat twisted, with the tusk alveoli (sockets) being divergent from each other at least in Pleistocene species.[10] These tusks could reach 4 metres (13 ft) in length, and probably over 190 kilograms (420 lb) in weight in the largest species, larger than any recorded in modern elephants.[11]
The molar teeth of Palaeoloxodon species typically show a "dot-dash-dot" wear pattern,[12] with the enamel folds concentrated into a major central structure at the midline of the tooth, which are flanked by smaller folds on either side, and the crowns of the tooth are generally proportionally narrow.[13] While during the early evolution of the genus, the molar teeth had a low number of lamellae and were brachydont (low crowned), in Pleistocene species the teeth became very hypsodont (high crowned) and increased the number of lamellae, though the lamellae frequency is distinctly lower than that reached by advanced mammoth species.[10]
Species of Palaeoloxodon varied widely in size. Large bulls of Palaeoloxodon recki, Palaeoloxodon antiquus and Palaeoloxodon namadicus are thought to have been over 4 metres (13 ft) tall at the shoulder and greater than 12 tons in body mass, exceeding the size of the largest known African bush elephants. In a 2015 study, one fragmentary unlocated femur of P. namadicus described in the 19th century was estimated to have belonged to an individual 5.2 metres (17 ft) tall and 22 tonnes in weight, exceeding the estimates for the otherwise largest known land mammals, the paraceratheres. However, this estimate is highly speculative and the author suggested that it should be "taken with a grain of salt".[2] In contrast, some of the island dwarf species are the smallest elephants known. The smallest species, P. cypriotes and P. falconeri, were only 1 metre (3.3 ft) tall as fully grown adults,[14][15] with the bulls of P. falconeri only having an estimated body mass of 250 kg (550 lb).[15]
 Life restoration of Palaeoloxodon namadicus
Life restoration of Palaeoloxodon namadicus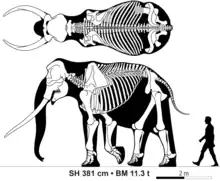 Skeletal diagram of an adult male Palaeoloxodon antiquus
Skeletal diagram of an adult male Palaeoloxodon antiquus.png.webp) Third molar teeth of Palaeoloxodon huaihoensis
Third molar teeth of Palaeoloxodon huaihoensis_(cropped).jpg.webp) Partial skull of Palaeoloxodon namadicus, showing the parieto-occipital crest at the top of the skull
Partial skull of Palaeoloxodon namadicus, showing the parieto-occipital crest at the top of the skull Size comparison of the dwarf elephant Palaeoloxodon falconeri, one of the smallest elephants known
Size comparison of the dwarf elephant Palaeoloxodon falconeri, one of the smallest elephants known Skull of Palaeoloxodon antiquus in front-on view, showing flared premaxillae with divergent tusks
Skull of Palaeoloxodon antiquus in front-on view, showing flared premaxillae with divergent tusks
Ecology
Species of Palaeoloxodon are thought to have similar social behaviour to modern elephants, with herds of adult females and juveniles, as well as solitary adult males.[16] The African species of Palaeoloxodon, as well as P. namadicus are suggested to have been grazers,[17][18] while P. antiquus is suggested to have been a variable mixed feeder than consumed a considerable amount of browse.[19]
Evolution
Palaeoloxodon first unambiguously appears in the fossil record in Africa during the Early Pleistocene, around 1.8 million years ago as the species Palaeoloxodon recki ileretensis (it is contested whether earlier "E. recki" subspecies are related to Palaeoloxodon).[20] P. recki was the dominant elephant in East Africa for most of the Pleistocene. A population of P. recki migrated out of Africa at the beginning of the Middle Pleistocene around 0.8 and 0.6 Mya, diversifying into the radiation of Eurasian Palaeoloxodon species, including P. antiquus, and P. namadicus. The precise relationships of the Eurasian taxa to each other are obscure.[21] The arrival of P. antiquus in Europe co-incides with the extinction of Mammuthus meridionalis and its replacement by Mammuthus trogontherii, suggesting that it might have shared a similar dietary niche and outcompeted the former.[21] P. antiquus was able to disperse onto many islands in the Mediterranean, undergoing insular dwarfism and speciating into numerous distinct varieties of dwarf elephants. Palaeoloxodon fossils are abundant in China and are assigned to three species, P. namadicus, P. naumanni and P. huaihoensis.[22] However, the relationships of Chinese Palaeoloxodon are currently unresolved and it is unclear how many species were present in the region.[9]
Extinction
The last Paleoloxodon species in Africa, P. iolensis, is suggested to have become extinct at the end of the Middle Pleistocene, around 130,000 years ago.[23] Most Eurasian species of Palaeoloxodon became extinct towards the end of the last glacial period. The youngest record of P. antiquus are footprints from the southern Iberian Peninsula, dating to approximately 28,000 years ago.[24] The youngest Japanese records of P. naumanni date to around 24,000 years ago.[25] The timing of extinction of Chinese Palaeoloxodon and Indian P. namadicus is uncertain, but claims of a Holocene survival are not substantiated for either region.[26][27][28] The youngest dates for the Sicilian dwarf elephant P. cf. mnaidriensis date to 32-20,000 years ago,[29] while the youngest dates for the Cyprus dwarf elephant P. cypriotes are around 12,000 years ago.[30] P. tiliensis from the Greek island of Tilos was suggested to have survived as recently as 3,500 years Before Present based on preliminary radiocarbon dating done in the 1970s, which would make it the youngest surviving elephant in Europe, but this has not been thoroughly investigated.[31]
Relationship with humans
Remains of the species P. recki, P. antiquus and P. naumanni have been found associated at a number of sites with stone stools and/or with cut marks on their bones, indicating that they were butchered, with some sites presenting clear evidence of hunting. These sites span from the Early Pleistocene, at least 1.3 but possibly as early as 1.6 million years ago to around 40,000 years ago, with the makers of the sites including both archaic humans and modern humans.[32][33][34]
References
- 1 2 Matsumoto, Hikoshichiro (1924). 日本産化石象の種類(略報) [Types of fossil elephants from Japan]. 地質学雑誌 (in Japanese). 31 (371): 255–272. doi:10.5575/geosoc.31.371_255.
- 1 2 Larramendi, A. (2015). "Shoulder height, body mass and shape of proboscideans". Acta Palaeontologica Polonica. 60. doi:10.4202/app.00136.2014.
- ↑ Shoshani, J.; Ferretti, M. P.; Lister, A. M.; Agenbroad, L. D.; Saegusa, H.; Mol, D.; Takahashi, K. (2007). "Relationships within the Elephantinae using hyoid characters". Quaternary International. 169–170: 174–185. Bibcode:2007QuInt.169..174S. doi:10.1016/j.quaint.2007.02.003.
- ↑ Meyer, Matthias; Palkopoulou, Eleftheria; Baleka, Sina; Stiller, Mathias; Penkman, Kirsty E H; Alt, Kurt W; Ishida, Yasuko; Mania, Dietrich; Mallick, Swapan; Meijer, Tom; Meller, Harald; Nagel, Sarah; Nickel, Birgit; Ostritz, Sven; Rohland, Nadin (2017-06-06). "Palaeogenomes of Eurasian straight-tusked elephants challenge the current view of elephant evolution". eLife. 6. doi:10.7554/eLife.25413. ISSN 2050-084X. PMC 5461109. PMID 28585920.
- ↑ Palkopoulou, Eleftheria; Lipson, Mark; Mallick, Swapan; Nielsen, Svend; Rohland, Nadin; Baleka, Sina; Karpinski, Emil; Ivancevic, Atma M.; To, Thu-Hien; Kortschak, R. Daniel; Raison, Joy M. (2018-03-13). "A comprehensive genomic history of extinct and living elephants". Proceedings of the National Academy of Sciences. 115 (11): E2566–E2574. Bibcode:2018PNAS..115E2566P. doi:10.1073/pnas.1720554115. ISSN 0027-8424. PMC 5856550. PMID 29483247.
- 1 2 Lin, Haifeng; Hu, Jiaming; Baleka, Sina; Yuan, Junxia; Chen, Xi; Xiao, Bo; Song, Shiwen; Du, Zhicheng; Lai, Xulong; Hofreiter, Michael; Sheng, Guilian (July 2023). "A genetic glimpse of the Chinese straight-tusked elephants". Biology Letters. 19 (7). doi:10.1098/rsbl.2023.0078. ISSN 1744-957X. PMC 10353889. PMID 37463654.
- ↑ Kevrekidis, C., & Mol, D. (2016). A new partial skeleton of Elephas (Palaeoloxodon) antiquus Falconer and Cautley, 1847 (Proboscidea, Elephantidae) from Amyntaio, Macedonia, Greece. Quaternary International, 406, 35–56. https://doi.org/10.1016/j.quaint.2015.11.110
- ↑ van der Geer, A.; Lyras, G. A.; de Vos, J. (2021). "Japan: Honshu, Shikoku, and Kyushu". Evolution of Island Mammals: Adaptation and Extinction of Placental Mammals on Islands. Oxford: Wiley Blackwell. p. 334. ISBN 9781119675730.
- 1 2 3 4 Larramendi, Asier; Zhang, Hanwen; Palombo, Maria Rita; Ferretti, Marco P. (February 2020). "The evolution of Palaeoloxodon skull structure: Disentangling phylogenetic, sexually dimorphic, ontogenetic, and allometric morphological signals". Quaternary Science Reviews. 229: 106090. Bibcode:2020QSRv..22906090L. doi:10.1016/j.quascirev.2019.106090. S2CID 213676377.
- 1 2 3 Athanassiou, Athanassios (2022), Vlachos, Evangelos (ed.), "The Fossil Record of Continental Elephants and Mammoths (Mammalia: Proboscidea: Elephantidae) in Greece", Fossil Vertebrates of Greece Vol. 1, Cham: Springer International Publishing, pp. 345–391, doi:10.1007/978-3-030-68398-6_13, ISBN 978-3-030-68397-9, S2CID 245067102, retrieved 2023-07-16
- ↑ Larramendi, Asier (2023-12-10). "Estimating tusk masses in proboscideans: a comprehensive analysis and predictive model". Historical Biology: 1–14. doi:10.1080/08912963.2023.2286272. ISSN 0891-2963.
- ↑ Pokines, James T.; Lister, Adrian M.; Ames, Christopher J. H.; Nowell, April; Cordova, Carlos E. (March 2019). "Faunal remains from recent excavations at Shishan Marsh 1 (SM1), a Late Lower Paleolithic open-air site in the Azraq Basin, Jordan". Quaternary Research. 91 (2): 768–791. Bibcode:2019QuRes..91..768P. doi:10.1017/qua.2018.113. ISSN 0033-5894. S2CID 134216492.
- ↑ Lister, Adrian M.; Dirks, Wendy; Assaf, Amnon; Chazan, Michael; Goldberg, Paul; Applbaum, Yaakov H.; Greenbaum, Nathalie; Horwitz, Liora Kolska (September 2013). "New fossil remains of Elephas from the southern Levant: Implications for the evolutionary history of the Asian elephant". Palaeogeography, Palaeoclimatology, Palaeoecology. 386: 119–130. Bibcode:2013PPP...386..119L. doi:10.1016/j.palaeo.2013.05.013. ISSN 0031-0182.
- ↑ Scarborough, Matthew Edward (2022-03-11). "Extreme Body Size Variation in Pleistocene Dwarf Elephants from the Siculo-Maltese Palaeoarchipelago: Disentangling the Causes in Time and Space". Quaternary. 5 (1): 17. doi:10.3390/quat5010017. hdl:11427/36354. ISSN 2571-550X.
- 1 2 Romano, Marco; Manucci, Fabio; Palombo, Maria Rita (2021-03-04). "The smallest of the largest: new volumetric body mass estimate and in-vivo restoration of the dwarf elephant Palaeoloxodon ex gr. P. falconeri from Spinagallo Cave (Sicily)". Historical Biology. 33 (3): 340–353. Bibcode:2021HBio...33..340R. doi:10.1080/08912963.2019.1617289. ISSN 0891-2963. S2CID 181855906.
- ↑ Neto de Carvalho, Carlos; Belaústegui, Zain; Toscano, Antonio; Muñiz, Fernando; Belo, João; Galán, Jose María; Gómez, Paula; Cáceres, Luis M.; Rodríguez-Vidal, Joaquín; Cunha, Pedro Proença; Cachão, Mario; Ruiz, Francisco; Ramirez-Cruzado, Samuel; Giles-Guzmán, Francisco; Finlayson, Geraldine (2021-09-16). "First tracks of newborn straight-tusked elephants (Palaeoloxodon antiquus)". Scientific Reports. 11 (1): 17311. Bibcode:2021NatSR..1117311N. doi:10.1038/s41598-021-96754-1. ISSN 2045-2322. PMC 8445925. PMID 34531420.
- ↑ Manthi, Fredrick Kyalo; Sanders, William J.; Plavcan, J. Michael; Cerling, Thure E.; Brown, Francis H. (September 2020). "Late Middle Pleistocene Elephants from Natodomeri, Kenya and the Disappearance of Elephas (Proboscidea, Mammalia) in Africa". Journal of Mammalian Evolution. 27 (3): 483–495. doi:10.1007/s10914-019-09474-9. ISSN 1064-7554. S2CID 198190671.
- ↑ Patnaik, Rajeev; Singh, Ningthoujam Premjit; Paul, Debajyoti; Sukumar, Raman (2019-11-15). "Dietary and habitat shifts in relation to climate of Neogene-Quaternary proboscideans and associated mammals of the Indian subcontinent". Quaternary Science Reviews. 224: 105968. Bibcode:2019QSRv..22405968P. doi:10.1016/j.quascirev.2019.105968. ISSN 0277-3791. S2CID 210307849.
- ↑ Rivals, Florent; Semprebon, Gina M.; Lister, Adrian M. (September 2019). "Feeding traits and dietary variation in Pleistocene proboscideans: A tooth microwear review". Quaternary Science Reviews. 219: 145–153. Bibcode:2019QSRv..219..145R. doi:10.1016/j.quascirev.2019.06.027. S2CID 200073388.
- ↑ Sanders, William J. (2023-07-07). Evolution and Fossil Record of African Proboscidea (1 ed.). Boca Raton: CRC Press. pp. 267–293. doi:10.1201/b20016. ISBN 978-1-315-11891-8.
- 1 2 Lister, Adrian M. (2004), "Ecological Interactions of Elephantids in Pleistocene Eurasia", Human Paleoecology in the Levantine Corridor, Oxbow Books, pp. 53–60, ISBN 978-1-78570-965-4, retrieved 2020-04-14
- ↑ Kang, Jia-Cih; Lin, Chien-Hsiang; Chang, Chun-Hsiang (2021-04-14). "Age and growth of Palaeoloxodon huaihoensis from Penghu Channel, Taiwan: significance of their age distribution based on fossils". PeerJ. 9: e11236. doi:10.7717/peerj.11236. ISSN 2167-8359. PMC 8052959. PMID 33954049.
- ↑ Manthi, Fredrick Kyalo; Sanders, William J.; Plavcan, J. Michael; Cerling, Thure E.; Brown, Francis H. (September 2020). "Late Middle Pleistocene Elephants from Natodomeri, Kenya and the Disappearance of Elephas (Proboscidea, Mammalia) in Africa". Journal of Mammalian Evolution. 27 (3): 483–495. doi:10.1007/s10914-019-09474-9. ISSN 1064-7554. S2CID 198190671.
- ↑ de Carvalho, Carlos Neto; Figueiredo, Silvério; Muniz, Fernando; Belo, João; Cunha, Pedro P.; Baucon, Andrea; Cáceres, Luis M.; Rodriguez-Vidal, Joaquín (2020-07-02). "Tracking the last elephants in Europe during the Würm Pleniglacial: the importance of the Late Pleistocene aeolianite record in SW Iberia". Ichnos. 27 (3): 352–360. Bibcode:2020Ichno..27..352D. doi:10.1080/10420940.2020.1744586. ISSN 1042-0940. S2CID 216504699.
- ↑ Iwase, Akira; Hashizume, Jun; Izuho, Masami; Takahashi, Keiichi; Sato, Hiroyuki (March 2012). "Timing of megafaunal extinction in the late Late Pleistocene on the Japanese Archipelago". Quaternary International. 255: 114–124. Bibcode:2012QuInt.255..114I. doi:10.1016/j.quaint.2011.03.029.
- ↑ Jukar, A.M.; Lyons, S.K.; Wagner, P.J.; Uhen, M.D. (January 2021). "Late Quaternary extinctions in the Indian Subcontinent". Palaeogeography, Palaeoclimatology, Palaeoecology. 562: 110137. Bibcode:2021PPP...56210137J. doi:10.1016/j.palaeo.2020.110137. S2CID 228877664.
- ↑ Turvey, Samuel T.; Sathe, Vijay; Crees, Jennifer J.; Jukar, Advait M.; Chakraborty, Prateek; Lister, Adrian M. (January 2021). "Late Quaternary megafaunal extinctions in India: How much do we know?". Quaternary Science Reviews. 252: 106740. Bibcode:2021QSRv..25206740T. doi:10.1016/j.quascirev.2020.106740. S2CID 234265221.
- ↑ Turvey, Samuel T.; Tong, Haowen; Stuart, Anthony J.; Lister, Adrian M. (September 2013). "Holocene survival of Late Pleistocene megafauna in China: a critical review of the evidence". Quaternary Science Reviews. 76: 156–166. Bibcode:2013QSRv...76..156T. doi:10.1016/j.quascirev.2013.06.030. ISSN 0277-3791.
- ↑ Palombo, Maria Rita; Antonioli, Fabrizio; Di Patti, Carolina; Valeria, Lo Presti; Scarborough, Matthew E. (2021-10-03). "Was the dwarfed Palaeoloxodon from Favignana Island the last endemic Pleistocene elephant from the western Mediterranean islands?". Historical Biology. 33 (10): 2116–2134. Bibcode:2021HBio...33.2116P. doi:10.1080/08912963.2020.1772251. ISSN 0891-2963. S2CID 225710152.
- ↑ Athanassiou, Athanassios; Herridge, Victoria; Reese, David S.; Iliopoulos, George; Roussiakis, Socrates; Mitsopoulou, Vassiliki; Tsiolakis, Efthymios; Theodorou, George (August 2015). "Cranial evidence for the presence of a second endemic elephant species on Cyprus". Quaternary International. 379: 47–57. Bibcode:2015QuInt.379...47A. doi:10.1016/j.quaint.2015.05.065. ISSN 1040-6182.
- ↑ Athanassiou, Athanassios; van der Geer, Alexandra A.E.; Lyras, George A. (August 2019). "Pleistocene insular Proboscidea of the Eastern Mediterranean: A review and update". Quaternary Science Reviews. 218: 306–321. Bibcode:2019QSRv..218..306A. doi:10.1016/j.quascirev.2019.06.028. ISSN 0277-3791. S2CID 199107354.
- ↑ Haynes, Gary (March 2022). "Late Quaternary Proboscidean Sites in Africa and Eurasia with Possible or Probable Evidence for Hominin Involvement". Quaternary. 5 (1): 18. doi:10.3390/quat5010018. ISSN 2571-550X.
- ↑ Konidaris, George E.; Tourloukis, Vangelis (2021-04-14). "Proboscidea-Homo interactions in open-air localities during the Early and Middle Pleistocene of western Eurasia: a palaeontological and archaeolocigal perspective". Human-Elephant Interactions: From Past to Present. doi:10.15496/publikation-55599.
- ↑ Kondo, Y.; Takeshita, Y.; Watanabe, T.; Seki, M.; Nojiri-ko Excavation Research Group (April 2018). "Geology and Quaternary Environments of the Tategahana Paleolithic Site in Nojiri-ko (Lake Nojiri), Nagano, Central Japan". Quaternary International. 471: 385–395. Bibcode:2018QuInt.471..385K. doi:10.1016/j.quaint.2017.12.012.
Genera of the order Proboscidea | ||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ||||||||||||||||||||||||||||||
| 
 | |||||||||||||||||||||||||||||
