



Regulatory-associated protein of mTOR also known as raptor or KIAA1303 is an adapter protein that is encoded in humans by the RPTOR gene.[5][6][7] Two mRNAs from the gene have been identified that encode proteins of 1335 (isoform 1) and 1177 (isoform 2) amino acids long.
Gene and expression
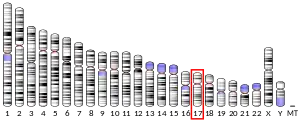
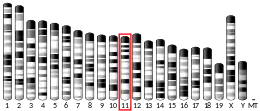
The human gene is located on human chromosome 17 with location of the cytogenic band at 17q25.3.[7]
Location
RPTOR is highly expressed in skeletal muscle and is somewhat less present in brain, lung, small intestine, kidney, and placenta tissue. Isoform 3 is widely expressed and most highly expressed in the nasal mucosa and pituitary. The lowest levels occur in the spleen.[8] In the cell, RPTOR is present in cytoplasm, lysosomes, and cytoplasmic granules. Amino acid availability determines RPTOR targeting to lysosomes. In stressed cells, RPTOR associates with SPAG5 and accumulates in stress granules, which significantly reduces its presence in lysosomes.[9][10]
Function
RPTOR encodes part of a signaling pathway regulating cell growth which responds to nutrient and insulin levels. RPTOR is an evolutionarily conserved protein with multiple roles in the mTOR pathway. The adapter protein and mTOR kinase form a stoichiometric complex. The encoded protein also associates with eukaryotic initiation factor 4E-binding protein-1 and ribosomal protein S6 kinase. It upregulates S6 kinase, the downstream effector ribosomal protein, and it downregulates the mTOR kinase. RPTOR also has a positive role in maintaining cell size and mTOR protein expression. The association of mTOR and RPTOR is stabilized by nutrient deprivation and other conditions which suppress the mTOR pathway.[8] Multiple transcript variants exist for this gene which encode different isoforms.[7]
Structure
RPTOR is a 150 kDa mTOR binding protein that is part of the mammalian target of rapamycin complex 1 (mTORC1). This complex contains mTOR, MLST8, RPTOR, AKT1S1/PRAS40, and DEPTOR. mTORC1 both binds to and is inhibited by FKBP12-rapamycin. mTORC1 activity is upregulated by mTOR and MPAK8 by insulin-stimulated phosphorylation at Ser-863.[11][12] MAPK8 also causes phosphorylation at Ser-696, Thr-706, and Ser-863 as a result of osmotic stress.[13] AMPK causes phosphorylation in the event of nutrient starvation and promotes 14-3-3 binding to raptor, which downregulates the mTORC1 complex.[14] RPS6KA1 stimulates mTORC1 activity by phosphorylating at Ser-719, Ser-721, and Ser-722 as a response to growth factors.
Interactions
- mTORC1 binds to and is inhibited by FKBP12-rapamycin
- RPTOR binds to 4EBP1 and RPS6KB1 directly whether or not it is associated with mTOR[15]
- RPTOR binds to poorly phosphorylated or non-phosphorylated EIF4EBP1 preferentially, which is important for mTOR to be able to catalyze phosphorylation.[6][15][16][17][18][19][20][21]
- RPTOR interacts with ULK1. This interaction depends on nutrients and is reduced in the case of starvation.[22]
- When RPTOR is phosphorylated by AMPK, it interacts with 14-3-3 protein and inhibits its activity.[14]
- RPTOR interacts with SPAG5, which competes with mTOR for binding RPTOR and causes decreased mTORC1 formation.
- RPTOR interacts with G3BP1. Oxidative stress increases the formation of the complex formed with RPTOR, G3BP1, and SPAG5[10]
RPTOR has also been shown to interact with:
Clinical significance
Signaling in cancer
The clinical significance of RPTOR is primarily due to its involvement in the mTOR pathway, which plays roles in mRNA translation, autophagy, and cell growth. Mutations in the PTEN tumor suppressor gene are the best known genetic deficiencies in cancer which affect mTOR signaling. These mutations are frequently found in a very large variety of cancers, including prostate, breast, lung, bladder, melanoma, endometrial, thyroid, brain, and renal carcinomas. PTEN inhibits the lipid-kinase activity of class I PtdIns3Ks, which phosphorylate PtdIns(4,5)P2 to create PtdIns(3,4,5)P3 (PIP3). PIP3 is a membrane-docking site for AKT and PDK1. In turn, active PDK1, along with mTORC1, phosphorylates S6K in the part of the mTOR pathway which promotes protein synthesis and cell growth.[39]
The mTOR pathway has also been found to be involved in aging. Studies with C. elegans, fruitflies, and mice have shown that the lifespan of the organism is significantly increased by inhibiting mTORC1.[40][41] mTORC1 phosphorylates Atg13 and stops it from forming the ULK1 kinase complex. This inhibits autophagy, the major degradation pathway in eukaryotic cells.[42] Because mTORC1 inhibits autophagy and stimulates cell growth, it can cause damaged proteins and cell structures to accumulate. For this reason, dysfunction in the process of autophagy can contribute to several diseases, including cancer.[43]
The mTOR pathway is important in many cancers. In cancer cells, astrin is required to suppress apoptosis during stress. Astrin recruits RPTOR to stress granules, inhibiting mTORC1 association and preventing apoptosis induced by mTORC1 hyperactivation. Because astrin is frequently upregulated in tumors, it is a potential target to sensitize tumors to apoptosis through the mTORC1 pathway.[10]
RPTOR is overexpressed in pituitary adenoma, and its expression increases with tumor staging. RPTOR could be valuable in the prediction and prognosis of pituitary adenoma due to this correlation between protein expression and the growth and invasion of the tumor.[44]
As a drug target
mTOR is found in two different complexes. When it associates with rapamycin-insensitive companion of mTOR (rictor), the complex is known as mTORC2 and it is insensitive to rapamycin. However, the complex mTORC1 formed by association with accessory protein RPTOR is sensitive to rapamycin. Rapamycin is a macrolide which is an immunosuppressant in humans that inhibits mTOR by binding to its intracellular receptor FKBP12. In many cancers, hyperactive AKT signaling leads to increased mTOR signaling, so rapamycin has been considered as an anti-cancer therapeutic for cancers with PTEN inactivation. Numerous clinical trials involving rapamycin analogs, such as CCI-779, RAD001, and AP23573, are ongoing. Early reports have been promising for renal-cell carcinoma, breast carcinomas, and non-small-cell lung carcinomas.[39]
See also
References
- 1 2 3 GRCh38: Ensembl release 89: ENSG00000141564 - Ensembl, May 2017
- 1 2 3 GRCm38: Ensembl release 89: ENSMUSG00000025583 - Ensembl, May 2017
- ↑ "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- ↑ "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- ↑ Nagase T, Kikuno R, Ishikawa KI, Hirosawa M, Ohara O (Apr 2000). "Prediction of the coding sequences of unidentified human genes. XVI. The complete sequences of 150 new cDNA clones from brain which code for large proteins in vitro". DNA Res. 7 (1): 65–73. doi:10.1093/dnares/7.1.65. PMID 10718198.
- 1 2 3 4 Hara K, Maruki Y, Long X, Yoshino K, Oshiro N, Hidayat S, Tokunaga C, Avruch J, Yonezawa K (Aug 2002). "Raptor, a binding partner of target of rapamycin (TOR), mediates TOR action". Cell. 110 (2): 177–89. doi:10.1016/S0092-8674(02)00833-4. PMID 12150926.
- 1 2 3 "Entrez Gene: KIAA1303 raptor".
- 1 2 3 Kim DH, Sarbassov DD, Ali SM, King JE, Latek RR, Erdjument-Bromage H, Tempst P, Sabatini DM (2002). "mTOR interacts with raptor to form a nutrient-sensitive complex that signals to the cell growth machinery". Cell. 110 (2): 163–75. doi:10.1016/S0092-8674(02)00808-5. PMID 12150925.
- ↑ Sancak Y, Bar-Peled L, Zoncu R, Markhard AL, Nada S, Sabatini DM (2010). "Ragulator-Rag complex targets mTORC1 to the lysosomal surface and is necessary for its activation by amino acids". Cell. 141 (2): 290–303. doi:10.1016/j.cell.2010.02.024. PMC 3024592. PMID 20381137.
- 1 2 3 Thedieck K, Holzwarth B, Prentzell MT, Boehlke C, Kläsener K, Ruf S, Sonntag AG, Maerz L, Grellscheid SN, Kremmer E, Nitschke R, Kuehn EW, Jonker JW, Groen AK, Reth M, Hall MN, Baumeister R (2013). "Inhibition of mTORC1 by astrin and stress granules prevents apoptosis in cancer cells". Cell. 154 (4): 859–74. doi:10.1016/j.cell.2013.07.031. PMID 23953116.
- ↑ Foster KG, Acosta-Jaquez HA, Romeo Y, Ekim B, Soliman GA, Carriere A, Roux PP, Ballif BA, Fingar DC (2010). "Regulation of mTOR complex 1 (mTORC1) by raptor Ser863 and multisite phosphorylation". J. Biol. Chem. 285 (1): 80–94. doi:10.1074/jbc.M109.029637. PMC 2804229. PMID 19864431.
- ↑ Carrière A, Cargnello M, Julien LA, Gao H, Bonneil E, Thibault P, Roux PP (2008). "Oncogenic MAPK signaling stimulates mTORC1 activity by promoting RSK-mediated raptor phosphorylation". Curr. Biol. 18 (17): 1269–77. doi:10.1016/j.cub.2008.07.078. PMID 18722121.
- ↑ Kwak D, Choi S, Jeong H, Jang JH, Lee Y, Jeon H, Lee MN, Noh J, Cho K, Yoo JS, Hwang D, Suh PG, Ryu SH (2012). "Osmotic stress regulates mammalian target of rapamycin (mTOR) complex 1 via c-Jun N-terminal Kinase (JNK)-mediated Raptor protein phosphorylation". J. Biol. Chem. 287 (22): 18398–407. doi:10.1074/jbc.M111.326538. PMC 3365776. PMID 22493283.
- 1 2 Gwinn DM, Shackelford DB, Egan DF, Mihaylova MM, Mery A, Vasquez DS, Turk BE, Shaw RJ (2008). "AMPK phosphorylation of raptor mediates a metabolic checkpoint". Mol. Cell. 30 (2): 214–26. doi:10.1016/j.molcel.2008.03.003. PMC 2674027. PMID 18439900.
- 1 2 3 Wang L, Rhodes CJ, Lawrence JC (2006). "Activation of mammalian target of rapamycin (mTOR) by insulin is associated with stimulation of 4EBP1 binding to dimeric mTOR complex 1". J. Biol. Chem. 281 (34): 24293–303. doi:10.1074/jbc.M603566200. PMID 16798736.
- 1 2 Schalm SS, Fingar DC, Sabatini DM, Blenis J (2003). "TOS motif-mediated raptor binding regulates 4E-BP1 multisite phosphorylation and function". Curr. Biol. 13 (10): 797–806. doi:10.1016/S0960-9822(03)00329-4. PMID 12747827.
- 1 2 3 Ha SH, Kim DH, Kim IS, Kim JH, Lee MN, Lee HJ, Kim JH, Jang SK, Suh PG, Ryu SH (2006). "PLD2 forms a functional complex with mTOR/raptor to transduce mitogenic signals". Cell. Signal. 18 (12): 2283–91. doi:10.1016/j.cellsig.2006.05.021. PMID 16837165.
- 1 2 3 Nojima H, Tokunaga C, Eguchi S, Oshiro N, Hidayat S, Yoshino K, Hara K, Tanaka N, Avruch J, Yonezawa K (2003). "The mammalian target of rapamycin (mTOR) partner, raptor, binds the mTOR substrates p70 S6 kinase and 4E-BP1 through their TOR signaling (TOS) motif". J. Biol. Chem. 278 (18): 15461–4. doi:10.1074/jbc.C200665200. PMID 12604610.
- ↑ Eguchi S, Tokunaga C, Hidayat S, Oshiro N, Yoshino K, Kikkawa U, Yonezawa K (2006). "Different roles for the TOS and RAIP motifs of the translational regulator protein 4E-BP1 in the association with raptor and phosphorylation by mTOR in the regulation of cell size". Genes Cells. 11 (7): 757–66. doi:10.1111/j.1365-2443.2006.00977.x. PMID 16824195. S2CID 30113895.
- ↑ Beugnet A, Wang X, Proud CG (2003). "Target of rapamycin (TOR)-signaling and RAIP motifs play distinct roles in the mammalian TOR-dependent phosphorylation of initiation factor 4E-binding protein 1". J. Biol. Chem. 278 (42): 40717–22. doi:10.1074/jbc.M308573200. PMID 12912989.
- ↑ Wang X, Beugnet A, Murakami M, Yamanaka S, Proud CG (2005). "Distinct signaling events downstream of mTOR cooperate to mediate the effects of amino acids and insulin on initiation factor 4E-binding proteins". Mol. Cell. Biol. 25 (7): 2558–72. doi:10.1128/MCB.25.7.2558-2572.2005. PMC 1061630. PMID 15767663.
- ↑ Hosokawa N, Hara T, Kaizuka T, Kishi C, Takamura A, Miura Y, Iemura S, Natsume T, Takehana K, Yamada N, Guan JL, Oshiro N, Mizushima N (2009). "Nutrient-dependent mTORC1 association with the ULK1-Atg13-FIP200 complex required for autophagy". Mol. Biol. Cell. 20 (7): 1981–91. doi:10.1091/mbc.E08-12-1248. PMC 2663915. PMID 19211835.
- 1 2 Jacinto E, Loewith R, Schmidt A, Lin S, Rüegg MA, Hall A, Hall MN (2004). "Mammalian TOR complex 2 controls the actin cytoskeleton and is rapamycin insensitive". Nat. Cell Biol. 6 (11): 1122–8. doi:10.1038/ncb1183. PMID 15467718. S2CID 13831153.
- 1 2 Sarbassov DD, Ali SM, Kim DH, Guertin DA, Latek RR, Erdjument-Bromage H, Tempst P, Sabatini DM (2004). "Rictor, a novel binding partner of mTOR, defines a rapamycin-insensitive and raptor-independent pathway that regulates the cytoskeleton". Curr. Biol. 14 (14): 1296–302. doi:10.1016/j.cub.2004.06.054. PMID 15268862.
- ↑ Ali SM, Sabatini DM (2005). "Structure of S6 kinase 1 determines whether raptor-mTOR or rictor-mTOR phosphorylates its hydrophobic motif site". J. Biol. Chem. 280 (20): 19445–8. doi:10.1074/jbc.C500125200. PMID 15809305.
- ↑ Long X, Lin Y, Ortiz-Vega S, Yonezawa K, Avruch J (2005). "Rheb binds and regulates the mTOR kinase". Curr. Biol. 15 (8): 702–13. doi:10.1016/j.cub.2005.02.053. PMID 15854902.
- 1 2 Jacinto E, Facchinetti V, Liu D, Soto N, Wei S, Jung SY, Huang Q, Qin J, Su B (2006). "SIN1/MIP1 maintains rictor-mTOR complex integrity and regulates Akt phosphorylation and substrate specificity". Cell. 127 (1): 125–37. doi:10.1016/j.cell.2006.08.033. PMID 16962653.
- ↑ Frias MA, Thoreen CC, Jaffe JD, Schroder W, Sculley T, Carr SA, Sabatini DM (2006). "mSin1 is necessary for Akt/PKB phosphorylation, and its isoforms define three distinct mTORC2s". Curr. Biol. 16 (18): 1865–70. doi:10.1016/j.cub.2006.08.001. PMID 16919458.
- ↑ Buerger C, DeVries B, Stambolic V (2006). "Localization of Rheb to the endomembrane is critical for its signaling function". Biochem. Biophys. Res. Commun. 344 (3): 869–80. doi:10.1016/j.bbrc.2006.03.220. PMID 16631613.
- ↑ McMahon LP, Yue W, Santen RJ, Lawrence JC (2005). "Farnesylthiosalicylic acid inhibits mammalian target of rapamycin (mTOR) activity both in cells and in vitro by promoting dissociation of the mTOR-raptor complex". Mol. Endocrinol. 19 (1): 175–83. doi:10.1210/me.2004-0305. PMID 15459249.
- ↑ Oshiro N, Yoshino K, Hidayat S, Tokunaga C, Hara K, Eguchi S, Avruch J, Yonezawa K (2004). "Dissociation of raptor from mTOR is a mechanism of rapamycin-induced inhibition of mTOR function". Genes Cells. 9 (4): 359–66. doi:10.1111/j.1356-9597.2004.00727.x. PMID 15066126. S2CID 24814691.
- ↑ Kawai S, Enzan H, Hayashi Y, Jin YL, Guo LM, Miyazaki E, Toi M, Kuroda N, Hiroi M, Saibara T, Nakayama H (2003). "Vinculin: a novel marker for quiescent and activated hepatic stellate cells in human and rat livers". Virchows Arch. 443 (1): 78–86. doi:10.1007/s00428-003-0804-4. PMID 12719976. S2CID 21552704.
- ↑ Choi KM, McMahon LP, Lawrence JC (2003). "Two motifs in the translational repressor PHAS-I required for efficient phosphorylation by mammalian target of rapamycin and for recognition by raptor". J. Biol. Chem. 278 (22): 19667–73. doi:10.1074/jbc.M301142200. PMID 12665511.
- ↑ Schieke SM, Phillips D, McCoy JP, Aponte AM, Shen RF, Balaban RS, Finkel T (2006). "The mammalian target of rapamycin (mTOR) pathway regulates mitochondrial oxygen consumption and oxidative capacity". J. Biol. Chem. 281 (37): 27643–52. doi:10.1074/jbc.M603536200. PMID 16847060.
- ↑ Sarbassov DD, Ali SM, Sengupta S, Sheen JH, Hsu PP, Bagley AF, Markhard AL, Sabatini DM (2006). "Prolonged rapamycin treatment inhibits mTORC2 assembly and Akt/PKB". Mol. Cell. 22 (2): 159–68. doi:10.1016/j.molcel.2006.03.029. PMID 16603397.
- ↑ Tzatsos A, Kandror KV (2006). "Nutrients suppress phosphatidylinositol 3-kinase/Akt signaling via raptor-dependent mTOR-mediated insulin receptor substrate 1 phosphorylation". Mol. Cell. Biol. 26 (1): 63–76. doi:10.1128/MCB.26.1.63-76.2006. PMC 1317643. PMID 16354680.
- ↑ Sarbassov DD, Sabatini DM (2005). "Redox regulation of the nutrient-sensitive raptor-mTOR pathway and complex". J. Biol. Chem. 280 (47): 39505–9. doi:10.1074/jbc.M506096200. PMID 16183647.
- ↑ Yang Q, Inoki K, Ikenoue T, Guan KL (2006). "Identification of Sin1 as an essential TORC2 component required for complex formation and kinase activity". Genes Dev. 20 (20): 2820–32. doi:10.1101/gad.1461206. PMC 1619946. PMID 17043309.
- 1 2 Guertin DA, Sabatini DM (July 2007). "Defining the role of mTOR in cancer". Cancer Cell. 12 (1): 9–22. doi:10.1016/j.ccr.2007.05.008. PMID 17613433.
- ↑ Robida-Stubbs S, Glover-Cutter K, Lamming DW, Mizunuma M, Narasimhan SD, Neumann-Haefelin E, Sabatini DM, Blackwell TK (May 2012). "TOR signaling and rapamycin influence longevity by regulating SKN-1/Nrf and DAF-16/FoxO". Cell Metab. 15 (5): 713–24. doi:10.1016/j.cmet.2012.04.007. PMC 3348514. PMID 22560223.
- ↑ Harrison DE, Strong R, Sharp ZD, Nelson JF, Astle CM, Flurkey K, Nadon NL, Wilkinson JE, Frenkel K, Carter CS, Pahor M, Javors MA, Fernandez E, Miller RA (July 2009). "Rapamycin fed late in life extends lifespan in genetically heterogeneous mice". Nature. 460 (7253): 392–5. Bibcode:2009Natur.460..392H. doi:10.1038/nature08221. PMC 2786175. PMID 19587680.
- ↑ Pyo JO, Nah J, Jung YK (February 2012). "Molecules and their functions in autophagy". Exp. Mol. Med. 44 (2): 73–80. doi:10.3858/emm.2012.44.2.029. PMC 3296815. PMID 22257882.
- ↑ Codogno P, Meijer AJ (November 2005). "Autophagy and signaling: their role in cell survival and cell death". Cell Death Differ. 12 (Suppl 2): 1509–18. doi:10.1038/sj.cdd.4401751. PMID 16247498.
- ↑ Jia W, Sanders AJ, Jia G, Liu X, Lu R, Jiang WG (August 2013). "Expression of the mTOR pathway regulators in human pituitary adenomas indicates the clinical course". Anticancer Res. 33 (8): 3123–31. PMID 23898069.
Further reading
- Kim DH, Sarbassov DD, Ali SM, King JE, Latek RR, Erdjument-Bromage H, Tempst P, Sabatini DM (2002). "mTOR interacts with raptor to form a nutrient-sensitive complex that signals to the cell growth machinery". Cell. 110 (2): 163–75. doi:10.1016/S0092-8674(02)00808-5. PMID 12150925.
- Nojima H, Tokunaga C, Eguchi S, Oshiro N, Hidayat S, Yoshino K, Hara K, Tanaka N, Avruch J, Yonezawa K (2003). "The mammalian target of rapamycin (mTOR) partner, raptor, binds the mTOR substrates p70 S6 kinase and 4E-BP1 through their TOR signaling (TOS) motif". J. Biol. Chem. 278 (18): 15461–4. doi:10.1074/jbc.C200665200. PMID 12604610.
- Kim DH, Sarbassov DD, Ali SM, Latek RR, Guntur KV, Erdjument-Bromage H, Tempst P, Sabatini DM (2003). "GbetaL, a positive regulator of the rapamycin-sensitive pathway required for the nutrient-sensitive interaction between raptor and mTOR". Mol. Cell. 11 (4): 895–904. doi:10.1016/S1097-2765(03)00114-X. PMID 12718876.
- Oshiro N, Yoshino K, Hidayat S, Tokunaga C, Hara K, Eguchi S, Avruch J, Yonezawa K (2004). "Dissociation of raptor from mTOR is a mechanism of rapamycin-induced inhibition of mTOR function". Genes Cells. 9 (4): 359–66. doi:10.1111/j.1356-9597.2004.00727.x. PMID 15066126. S2CID 24814691.
- Sarbassov DD, Ali SM, Kim DH, Guertin DA, Latek RR, Erdjument-Bromage H, Tempst P, Sabatini DM (2004). "Rictor, a novel binding partner of mTOR, defines a rapamycin-insensitive and raptor-independent pathway that regulates the cytoskeleton". Curr. Biol. 14 (14): 1296–302. doi:10.1016/j.cub.2004.06.054. PMID 15268862.
- Beausoleil SA, Jedrychowski M, Schwartz D, Elias JE, Villén J, Li J, Cohn MA, Cantley LC, Gygi SP (2004). "Large-scale characterization of HeLa cell nuclear phosphoproteins". Proc. Natl. Acad. Sci. U.S.A. 101 (33): 12130–5. Bibcode:2004PNAS..10112130B. doi:10.1073/pnas.0404720101. PMC 514446. PMID 15302935.
- Jacinto E, Loewith R, Schmidt A, Lin S, Rüegg MA, Hall A, Hall MN (2004). "Mammalian TOR complex 2 controls the actin cytoskeleton and is rapamycin insensitive". Nat. Cell Biol. 6 (11): 1122–8. doi:10.1038/ncb1183. PMID 15467718. S2CID 13831153.
- Sarbassov DD, Guertin DA, Ali SM, Sabatini DM (2005). "Phosphorylation and regulation of Akt/PKB by the rictor-mTOR complex". Science. 307 (5712): 1098–101. Bibcode:2005Sci...307.1098S. doi:10.1126/science.1106148. PMID 15718470. S2CID 45837814.
- Long X, Lin Y, Ortiz-Vega S, Yonezawa K, Avruch J (2005). "Rheb binds and regulates the mTOR kinase". Curr. Biol. 15 (8): 702–13. doi:10.1016/j.cub.2005.02.053. PMID 15854902.
- Sarbassov DD, Sabatini DM (2005). "Redox regulation of the nutrient-sensitive raptor-mTOR pathway and complex". J. Biol. Chem. 280 (47): 39505–9. doi:10.1074/jbc.M506096200. PMID 16183647.
- Tzatsos A, Kandror KV (2006). "Nutrients suppress phosphatidylinositol 3-kinase/Akt signaling via raptor-dependent mTOR-mediated insulin receptor substrate 1 phosphorylation". Mol. Cell. Biol. 26 (1): 63–76. doi:10.1128/MCB.26.1.63-76.2006. PMC 1317643. PMID 16354680.
- Shah OJ, Hunter T (2006). "Turnover of the active fraction of IRS1 involves raptor-mTOR- and S6K1-dependent serine phosphorylation in cell culture models of tuberous sclerosis". Mol. Cell. Biol. 26 (17): 6425–34. doi:10.1128/MCB.01254-05. PMC 1592824. PMID 16914728.
- Kudchodkar SB, Yu Y, Maguire TG, Alwine JC (2006). "Human cytomegalovirus infection alters the substrate specificities and rapamycin sensitivities of raptor- and rictor-containing complexes". Proc. Natl. Acad. Sci. U.S.A. 103 (38): 14182–7. Bibcode:2006PNAS..10314182K. doi:10.1073/pnas.0605825103. PMC 1599931. PMID 16959881.
- Beausoleil SA, Villén J, Gerber SA, Rush J, Gygi SP (2006). "A probability-based approach for high-throughput protein phosphorylation analysis and site localization". Nat. Biotechnol. 24 (10): 1285–92. doi:10.1038/nbt1240. PMID 16964243. S2CID 14294292.
- Olsen JV, Blagoev B, Gnad F, Macek B, Kumar C, Mortensen P, Mann M (2006). "Global, in vivo, and site-specific phosphorylation dynamics in signaling networks". Cell. 127 (3): 635–48. doi:10.1016/j.cell.2006.09.026. PMID 17081983.