
The timeline of therizinosaur research is a chronological listing of events in the history of paleontology focused on therizinosaurs. They were unusually long-necked, pot-bellied, and large-clawed herbivorous theropods most closely related to birds. The early history of therizinosaur research occurred in three phases. The first phase was the discovery of scanty and puzzling fossils in Asia by the Central Asiatic Expeditions of the 1920s and Soviet-backed research in the 1950s. This phase resulted in the discovery of the Therizinosaurus cheloniformis type specimen. Soviet paleontologist Evgeny Maleev interpreted these unusual remains as belonging to some kind of gigantic turtle.
The second major phase of therizinosaur research followed the discovery of better preserved remains in the 1970s by collaborative research between the Soviets and Mongolians. These finds revealed the true nature of therizinosaurs as bizarre dinosaurs. However, the exact nature and classification of therizinosaurs within Dinosauria was controversial as was their paleobiology. When Rozhdestventsky first reinterpreted therizinosaurs as dinosaurs he argued that they were unusual theropods that may have used their clawed arms to break open termite mounds or collect fruit. Osmolska and Roniewicz also considered therizinosaurs to be theropods.
In 1979, Altangerel Perle named the new species Segnosaurus galbinensis, which although he recognized was an unusual theropod, he did not recognize as a therizinosaur. Consequently, he named the new family Segnosauridae and, in 1980, Segnosauria. Two years later, Perle recognized commonalities between Therizinosaurus and segnosaurs, reclassifying the former as a member of the latter. From hereout therizinosaur research was considered "segnosaur" research. Perle himself thought that his "segnosaurs" were semi-aquatic fish-eaters. However, in the early 1990s, researchers like Rinchen Barsbold and Teresa Maryańska cast doubt on the connection between therizinosaurs and segnosaurs altogether.
Nevertheless, the description Alxasaurus elsitaiensis provided more evidence for a close relationship between the therizinosaurs and "segnosaurs" and led to a revision of their classification. The discovery of this and other primitive therizinosaurs in China formed the beginnings of the third major wave of therizinosaur research. That same year Russell and Russell reinterpreted therizinosaurs as herbivorous foragers like mammalian chalicotherium. Other significant finds of the 1990s include therizinosaur eggs with embryos preserved inside and the first known therizinosaur with feathers, Beipiaosaurus, which was described from China in 1999.
20th century
1950s

- Evgeny Maleev described the new genus and species Therizinosaurus cheloniformis. from the Nemegt Formation He interpreted it as a gigantic turtle. Maleev also named the family Therizinosauridae to contain this species.[1]
1960s
- Zakharov described and named the genus and type ichnospecies Macropodosaurus gravis, which is represented by a series of four-toed footprints.[2]
1970s

- Rozhdestventsky first proposed the idea that therizinosaurids were actually theropod dinosaurs. He thought they used their large claws to tear open termite mounds or collect fruit from trees.[3]
- Osmolska and Roniewicz also interpreted Therizinosaurus as a carnosaur theropod.[4]
- Barsbold proposed the Deinocheirosauria and included Therizinosaurus as a member.[5]
- In another paper during the same year, Barsbold referred a shoulder and forearm found in the same strata as the Therizinosaurus type specimen to that genus because of the resemblance between the specimens claws. He observed that the anatomy of the arm and shoulder remains suggested that it belonged to a theropod dinosaur. Barsbold also remarked on similarities it shared with Deinocheirus, another mysterious dinosaur from the same rock unit.[6]

- Dong described the new genus and species Nanshiungosaurus brevispinus based on a vertebral column and pelvis. He interpreted Nanshiungosaurus as a new genus of dwarf sauropods. He also described the new species Chilantaisaurus zheziangensis but interpreted it as a carnosaur.[7]
- Perle described the new genus and species Segnosaurus galbinensis based on mostly complete limbs and girdles. He erected a new family, the Segnosauridae, for this unusual dinosaur. He tentatively regarded it as a theropod.[8]
1980s

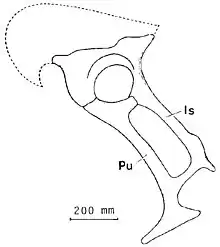


- Barsbold and Perle named the Segnosauria and described the new genus and species Erlikosaurus andrewsi. They also described in brief detail an unknown segnosaur. Barsbold and Perle thought segnosaurs were slow, semi-aquatic animals.[9]
- Perle redescribed the holotype of Erlikosaurus but this time in more detail and misspelled Erlicosaurus.[10]
- Perle reported an unusual partial four-toed hind limb from Hermiin Tsav, Nemegt Formation. Because this partial leg was found not far from where Barsbold reported the shoulder and humerus he referred to Therizinosaurus, Perle thought that his specimen also probably belonged to that taxon. Since this leg was similar to those of segnosaurs, he classified Therizinosaurus as a segnosaur.[11]
- Barsbold described the new genus and species Enigmosaurus mongoliensis based on a partial pelvis from the previously unknown segnosaur. Barsbold regarded Enigmosaurus as so unusual that he gave it its own family, the Enigmosauridae.[12]
- Paul suggested that segnosaurs shared an evolutionary relationship with prosauropods and ornithischians. He thought this implied that they were probably herbivores.[13]
- Gauthier considered segnosaurs to be relatives of sauropodomorphs.[14]
- Paul Sereno also followed this new interpretation of segnosaurs.[15]
- Bryn J. Mader and colleagues re-examined the specimens assigned to Alectrosaurus and concluded that the large humerus and unguals used by Gilmore in 1933 did not come from a tyrannosauroid, but a therizinosaurid.[16]
1990s
- Barsbold and Maryanska reinterpreted the sauropod Nanshiungosaurus and the carnosaur Chilantaisaurus as segnosaurs. They agreed with Perle that the partial hind limb from Hermiin Tsav he described in 1982 was segnosaurian, but cast doubt with his referral of it to Therizinosaurus, and therefore with his subsequent conclusion that Therizinosaurus was a segnosaur. Barsbold and Maryanska also disagreed with previous researchers who classified Deinocheirus as a segnosaur.[17]
- David B. Norman considered Therizinosaurus to be a theropod of uncertain classification.[18]
- A collaborative expedition between Chinese and Japanese scientists discovered the type specimen of the new segnosaur species. In addition, Dong named the Segnosaurischia to place segnosaurs on an equal rank with Saurischia and Ornithischia.[19]

- Russel and Dong described the new genus and species Alxasaurus elsitaiensis. They considered it distinct enough to warrant its own family, the Alxasauridae. They pointed out similar anatomical traits in the anatomy of the hand of the partial forelimb Barsbold referred to Therizinosaurus in 1976 and those of segnosaurs. They concluded that therizinosaurids and segnosaurids were theropods and almost identical in traits, and synonymized the Therizinosauridae and Segnosauridae, with the former having nomenclatural priority and coined the superfamily Therizinosauroidea to contain Alxasaurus and its family. The relative completeness of the specimens concluded the debate on whether or not segnosaurs were theropods in the affirmative.[20]
- Russell and Russell noticed that segnosaurs had similar body plans to chalicotheres and ground sloths and concluded that they may have used their large forelimbs to forage in a similar manner.[21]

- Clark and others redescribed the skull of Erlikosaurus. They found more evidence that segnosaurs were theropods and classified them as maniraptorans.[22]
- Nessov speculated that segnosaurs may have hung from trees using their large claws like sloths do and fed on wasp nests. He reported the discovery of segnosaur remains in Kazakhstan and Uzbekistan.[23]
- Currie attributed some Late Cretaceous eggs and their embryos from the Nanchao Formation of Henan, China to segnosaur dinosaurs. These spherical eggs still preserve the fossilized remains of their developing embryos.
- In a second paper, Currie attributed another kind of fossil egg from an entirely different oofamily to segnosaurs. Unlike the spherical eggs of his first paper, these huge "elongated" eggs are classified as members of the Elongatoolithidae and could reach lengths of up to 50 cm.
- Manning and others did not agree with Currie's referral of elongatoolithid eggs to segnosaurs.[24]
- Dong and Yu described the new species "Nanshiungosaurus bohlini" discovered during the 1992 Sino-Japanese expedition, and coined the Nanshiungosauridae to contain it and Nanshiungosaurus.[25]
- Russell coined the Therizinosauria in order to contain all segnosaurs. This new infraorder was composed of Therizinosauroidea and the more advanced Therizinosauridae. With this, the terms segnosaur and Segnosauria became synonyms to therizinosaur and Therizinosauria, respectively.[26]
- Zhao and Xu reported the existence of a possible therizinosaur dentary dating all the way back to the Early Jurassic.[27]

- Xu, Tang, and Wang described the new genus and species Beipiaosaurus inexpectus. The type specimen actually preserves impressions of the animal's feathered integument. It was also the oldest known therizinosaur.[28]
- Carpenter reported that the embryos Currie considered therizinosaurian had teeth in their premaxillae, unlike any known member of the group. This could be evidence that the egglayer was actually primitive for a therizinosaur. He also did not confirm Currie's referral of elongatoolithid eggs to therizinosaurs.[29]
21st century
2000s
- Manning and others observed that the embryos identified by Currie in 1996 as therizinosaurian had "an unusual pattern of tooth replacement in which a slender, elongate tooth is replaced by a symmetrical, denticulate tooth". Contrary to Carpenter's claim in 1999, Manning and his colleagues reported the embryos premaxillae as toothless.[30]


- Xu, Zhao, and Clark described the new genus and species Eshanosaurus deguchiianus based on the Early Jurassic dentary. This species was the probably oldest known therizinosaur.[31]
- Kirkland and Wolfe described the new genus and species Nothronychus mckinleyi. This was the first definitive therizinosaurid discovered outside of Asia. They also found that Eshanosaurus had a similar dentition to prosauropods.[32]
- Zhang and others described the new genus and species Neimongosaurus yangi.[33]
- Xu and others described the new genus and species Erliansaurus bellamanus.[34]
- Xu and team described new pelvic and tail material from the holotype of Beipiaosaurus. These were discovered during a re-excavation of the fossil quarry were the first elements of the holotype were found.[35]
- Kirkland and colleagues briefly reported and described primitive remains of therizinosaurs found in Utah.[36]
- The same team made comparisons with braincases found among the remains and that of Nothronychus mckinleyi.[37]
- Lindsay Zanno continue with these short descriptions but this time focused on forelimb elements.[38]
- Zanno discussed in more detail these elements in her Ph.D.[39]
- Kirkland and others described the primitive new genus and species Falcarius utahensis based on the multiple remains previously described.[40]
- Zanno and Erickson briefly discussed the growth of Falcarius which was represented by adult and juvenile specimens.[41]
- Zanno formally published and described the pectoral anatomy of Falcarius.[42]
- Bursh reported the inferred range of motion in the therizinosaurids Neimongosaurus, which was very pronounced and circular.[43]
- Andrey G. Sennikov re-examined Macropodosaurus and concluded that a therizinosaurid-grade dinosaur made those tracks, suggesting a possible plantigrade stance. He also considered these tracks to be more associated with therizinosaurids.[44]

- Li and others described the new genus and species Suzhousaurus megatherioides.[45]
- Kundrát with colleagues described in detail the embryos from the Nanchao Formation.[46]
- A new specimen of Suzhousaurus was described by Li and colleagues.[47]
- Lee and colleagues reported new therizinosaurid specimens composed of partially complete skeletons collected from the Nemegt Formation.[48]
- Zanno and colleagues described the newer N. graffami, a second and more complete species of the genus Nothronychus.[49]
- Barrett considered that the highly dubious and controversial Eshanosaurus was a therizinosaur taxon.[50]
- Xu and colleagues described a new specimen of Beipiaosaurus preserving a partial skeleton and extensive feather integument. The latter were so unique compared to other feather types that they had to describe a new, primitive type of feathers named EBFFs (elongated broad filamentous feathers).[51]
2010s

- Zanno described in detail the osteology of Falcarius based on numerous specimens.[52]
- Zanno conducted the most detailed phylogenetic analysis of the Therizinosauria to that point. She cited the inaccessibility, damage, potential loss of holotype specimens, scarcity of cranial remains, and fragmentary specimens with few overlapping elements as the most significant obstacles to resolving the evolutionary relationships within the group. She also revised Therizinosauroidea to exclude Falcarius and retained it in the wider clade Therizinosauria, which became the senior synonym of Segnosauria.[53]
- Senter and James suggested the ecological niches for Therizinosaurus and Deinocheirus were characterized by high-browsing life styles. Therizinosaurus was found to be among the tallest dinosaurs in the Nemegt Formation. They also calculated that an average Therizinosaurus may have had 3 m (9.8 ft) long hindlimbs.[54]
- Smith and team described in detail the braincase anatomy of Falcarius.[55]
- Zanno and Makovicky found therizinosaurs and other groups of herbivorous dinosaurs that had beaks and retained teeth were unable to lose their teeth completely because they lacked gastric mills (gizzards) and needed the teeth to process food. The high-fiber folivorous (leaf-based) diet of therizinosaurs and other archosaurs may also have precluded the evolution of a complete beak.[56]



- Qian and colleagues discussed the affinities of Chilantaisaurus zheziangensis and noted that is actually a therizinosaurid. Tiantaisaurus, a supposedly new therizinosaur, was briefly mentioned but not officially named.[57]
- Lautenschlager with colleagues reconstructed the cranial cavities of Erlikosaurus via CT scans and noted a complex ear and brain structure that may apply to other therizinosaurids.[58]
- Senter with others described the new genus and species Martharaptor greenriverensis.[59]
- Fiorillo and Adams described four-toed footprints from the Cantwell Formation. The morphology is similar to therizinosaurid feet but slightly different from Macropodosaurus. Nevertheless, they attributed these tracks to therizinosaurids.[60]
- Lautenschlager performed digital reconstructions for the cranial musculature in Erlikosaurus and found the bite force of Edmontosaurus being greater than that for the former. The lesser bite force for Erlikosaurus better served in stripping and cropping leaves, rather than active mastication.[61]
- Using the complete holotype skull of Erlikosaurus, Lautenschlager and colleagues noted that the keratinous beak in therizinosaurs and most other beaked theropods was an adaption that might have helped to enhance cranial stability by mitigating the stress and strain experienced by the skull during feeding.[62]
- Pu with colleagues described the new primitive genus and species Jianchangosaurus yixianensis. They regarded this genus along with Falcarius was the most primitive.[63]
- Kobayashi and colleagues reported an exceptional nesting ground site of therizinosaurid dinosaurs at the Javkhlant Formation, which contained at least 17 dendroolithid egg clutches.[64]
- Lautenschlager concluded that the claws of most therizinosaurs were more effective when piercing or pulling down vegetation but not for digging. He could neither confirm nor disregard that the hand claws could have been fully used for sexual display, self-defense, intraspecific competition, mate-gripping during mating or grasping stabilization when foraging.[65]
- Li and colleagues examined numerous dinosaur specimens with preserved integument, and inferred that the specimen BMNHC PH000911 of Beipiaosaurus had a brownish colouration in life.[66]
- The osteology and taphonomy of Nothronychus was fully described by Hedrick and colleagues, providing anatomical considerations for other therizinosaurids.[67]
- David K. Smith reconstructed the cranio-cervical musculature of Falcarius and Nothronychus.[68]
- Kobayashi and team reported a new, two-fingered therizinosaur from the Bayan Shireh Formation that was discovered back in 2012.[69]
- Gierlinski reported Macropodosaurus footprints from Late Cretaceous strata in Poland.[70]
- Zanno with colleagues redescribed the complex lower jaws and dentition of Segnosaurus and noticed a niche partitioning between this taxon and Erlikosaurus.[71]
- Sues and Averianov described extensive remains of therizinosauroid and potentially therizinosaurid remains from the Bissekty Formation.[72]
- Botelho and team considered the feet of therizinosaurids to be plantigrade.[73]
- Lautenschlager concluded the evolutionary trends in jaw mechanics of therizinosaurs suffered a change from higher bite forces and robust lower jaws in early members to lesser ones in derived therizinosaurs.[74]
- Masrour with colleagues reported Macropodosaurus footprints on Cretaceous strata in Morocco.[75]
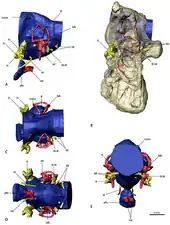
- Khai Button and team examined the dental histology of Falcarius and Suzhousaurus to analyze the dental evolution within Therizinosauria.[76]
- The holotype braincase of Nothronychus mckinleyi was re-analyzed by Smith with colleagues finding similar traits and capacities to Erlikosaurus.[77]
- Fiorillo and team found more therizinosaurid tracks from the Cantwell Formation but this time in association with hadrosaurid footprints. They were the first authors providing photographs of the holotype Erlikosaurus feet.[78]
- McNamara and colleagues discovered the fossilised remains of skin flakes in a specimen of Beipiaosaurus and other numerous feathered dinosaurs from the Jehol Biota using scanning electron microscope on the preserved feather impressions. Comparisons made with extant birds indicate that Beipiaosaurus and other non-avian theropods shed their skin as dandruff, and did not get as warm as modern birds because they were ground-dwelling animals not able to fly.[79]
- Loredana Macaluso and colleagues suggested that the pubic boot of most therizinosaurs was restrained by ventilatory muscles that were crucial for breathing with air sacs.[80]

- Hartman with colleagues performed a large phylogenetic analysis for the Therizinosauria and other theropod groups. This analysis was strongly based on the 2010 work of Zanno.[81]
- Yao and colleagues described the new genus and species Lingyuanosaurus sihedangensis.[82]
- Liao and Xu redescribed the holotype skull of Beipiaosaurus in detail, noting new unique cranial traits for the genus.[83]
- Ali Nabavizadeh concluded that most therizinosaurs were mainly orthal feeders (moving their jaws up and down and not to the sides) and raised their jaws isognathously whereby the upper and lower teeth of each side contacted each other at once.[84]
- Button and Zanno found that Segnosaurus had gracile skulls and relatively low bite forces, indicating a food processing in the gut. Erlikosaurus had features associated with extensive processing such as the lower jaws or dentition and therefore, the food processing was in the mouth.[85]
- The Javkhlant Formation nesting site was formally described in 2019 by Kohei Tanaka and colleagues concluding that egg clutches were covered in organic-rich material during incubation and colonial nesting first evolved in non-avian dinosaur to increase hatching success.[86]
2020s
- David K. Smith and colleagues analyzed the vertebral pneumaticity of Nothronychus concluding that it had bird-like unidirectional lungs and abdominal air sacs.[87]
.png.webp)
- Smith reconstructed the hindlimb musculature of Nothronychus, noting a convergence with birds and ornithischian dinosaurs.[88]
- Smith reconstructed the forelimb musculature of Nothronychus, discovering similarities with dromaeosaurid muscles and the absence of an increased range of motion within the forelimbs.[89]
- Chun-Chi Liao and colleagues redescribed the holotype of Beipiaosaurus, noting new unique traits for the genus.[90]
- Soki Hattori with colleagues redescribed the holotype of Fukuivenator considering numerous new traits of the genus. The team also recovered the genus as the primitive-most therizinosaur.[91]
- Kundrát and Arthur R. I. Cruickshankb presented evidence that some dendroolithid eggs were laid by a therizinosauroids.[92]
- Sennikov re-analyzed Macropodosaurus and the overall foot anatomy of therizinosaurids, concluding that this group had a plantigrade stance. While ancestral therizinosaurs had a gracile and digitigrade stance, therizinosaurids evolved a robust, ponderous body plan and plantigradism.[93]

- Kobayashi and team named and described the new therizinosaurid Paralitherizinosaurus japonicus from the Osoushinai Formation of Japan. It was mostly similar to Therizinosaurus in manual ungual morphology.[94]
- Zichuan Qin and team examined the manual unguals of therizinosaurs and alvarezsaurs through finite element analysis, concluding that the former group was best suited for herbivory. Special attention was given to Therizinosaurus, noting that its manual unguals became too enlarged to be mechanically functional.[95]
- Smith and David D. Gillette reconstructed the hindlimbs tendon/ligament system of Nothronychus, noting a slow, waddling gait. The co-workers also noted that a plantigrade stance in the group cannot be definitely rejected.[96]
- Smith reconstructed the hindlimb musculature of Falcarius, characterized by a non-waddling gait.[97]
- Simon Wills and colleagues reported the presence of a therizinosaur tooth from the Middle Jurassic Hornsleasow Quarry of the Chipping Norton Limestone Formation.[98]
See also
References
- ↑ Maleev, E. A. (1954). "Новый черепахообразный ящер в Монголии" [New turtle like reptile in Mongolia]. Priroda (3): 106−108. Translated paper
- ↑ Zakharov, S. A. (1964). "О сеноманском динозавре, следы которого обнаружены в долине р.Ширкент" [On the Cenomanian dinosaur, the tracks of which were found in the Shirkent River Valley]. In Reiman, V. M. (ed.). Paleontology of Tajikistan (in Russian). Dushanbe: Academy of Sciences of Tajik S.S.R. Press. pp. 31−35.
- ↑ Rozhdestvensky, A. K. (1970). "О гигантских когтевых фалангах загадочных рептилий мезозоя" [Giant claws of enigmatic Mesozoic reptiles]. Paleontological Journal (in Russian). 1970 (1): 131−141.
- ↑ Osmólska, H.; Roniewicz, E. (1970). "Deinocheiridae, a new family of theropod dinosaurs" (PDF). Palaeontologica Polonica (21): 5−19. Archived from the original (PDF) on 2016-03-03. Retrieved 2020-08-18.
- ↑ Barsbold, R. (1976). "On the evolution and systematics of the late Mesozoic dinosaurs". Trudy – Sovmestnaya Sovetsko-Mongol'skaya Paleontologicheskaya Ekspeditsiya (in Russian). 3: 68–75.
- ↑ Barsbold, R. (1976). "Новые данные о теризинозавре (Therizinosauridae, Theropoda)" [New data on Therizinosaurus (Therizinosauridae, Theropoda)]. In Kramarenko, N. N.; Luvsandansan, B.; Voronin, Y. I.; Barsbold, R.; Rozhdestvensky, A. K.; Trofimov, B. A.; Reshetov, V. Y. (eds.). Paleontology and Biostratigraphy of Mongolia. The Joint Soviet-Mongolian Paleontological Expedition, Transactions. Moscow: Nauka Press. pp. 76−92.
- ↑ Dong, Z. (1979). "Cretaceous dinosaur fossils in southern China" [Cretaceous dinosaurs of the Huanan (south China)]. In Institute of Vertebrate Paleontology and Paleoanthropology; Nanjing Institute of Paleontology (eds.). Mesozoic and Cenozoic Redbeds in Southern China (in Chinese). Beijing: Science Press. pp. 342−350. Translated paper
- ↑ Perle, A. (1979). "Segnosauridae - новое семейство позднемеловых хищных динозавров Монголии" [Segnosauridae - a new family of theropods from the Late Cretaceous of Mongolia]. Transactions of the Joint Soviet-Mongolian Paleontological Expedition (in Russian). 8: 45−55. Translated paper
- ↑ Barsbold, R.; Perle, A. (1980). "Segnosauria, a new suborder of carnivorous dinosaurs" (PDF). Acta Palaeontologica Polonica. 25 (2): 190−192.
- ↑ Perle, A. (1981). "Новый сегнозаврид из верхнего мела Монголии" [New segnosaurid from the Upper Cretaceous of Mongolia]. Transactions of the Joint Soviet-Mongolian Paleontological Expedition (in Russian). 15: 50−59. Translated paper
- ↑ Perle, A. (1982). "A hind limb of Therizinosaurus from the Upper Cretaceous of Mongolia". Problems in Mongolian Geology (in Russian). 5: 94−98. Translated paper
- ↑ Barsbold, R. (1983). "Хищные динозавры мела Монголии" [Carnivorous dinosaurs from the Cretaceous of Mongolia] (PDF). Transactions of the Joint Soviet-Mongolian Paleontological Expedition (in Russian). 19: 89. Translated paper
- ↑ Paul, G. S. (1984). "The segnosaurian dinosaurs: relics of the prosauropod-ornithischian transition?". Journal of Vertebrate Paleontology. 4 (4): 507−515. Bibcode:1984JVPal...4..507P. doi:10.1080/02724634.1984.10012026. ISSN 0272-4634. JSTOR 4523011.
- ↑ Gauthier, J. (1986). "Saurischian monophyly and the origin of birds". Memoirs of the California Academy of Sciences. 8: 45. Archived from the original on 2019-08-16.
- ↑ Sereno, P. (1989). "Prosauropod monophyly and basal sauropodomorph phylogeny". Abstract of Papers. Forty-Ninth Annual Meeting Society of Vertebrate Paleontology. Journal of Vertebrate Paleontology. Vol. 9, no. 3 Supplement. p. 39A. ISSN 0272-4634. JSTOR 4523276.
- ↑ Mader, B. J.; Bradley, R. L. (1989). "A redescription and revised diagnosis of the syntypes of the Mongolian tyrannosaur Alectrosaurus olseni". Journal of Vertebrate Paleontology. 9 (1): 41–55. Bibcode:1989JVPal...9...41M. doi:10.1080/02724634.1989.10011737.
- ↑ Barsbold, R.; Maryańska, T. (1990). "Saurischia Sedis Mutabilis: Segnosauria". In Weishampel, D. B.; Osmolska, H.; Dodson, P. (eds.). The Dinosauria (1st ed.). Berkeley: University of California Press. pp. 408−415. ISBN 9780520067271.
- ↑ Norman, D. B. (1990). "Problematic Theropoda: Coelurosauria". In Weishampel, D. B.; Osmolska, H.; Dodson, P. (eds.). The Dinosauria (1st ed.). Berkeley: University of California Press. pp. 280−305. ISBN 9780520067271.
- ↑ Dong, Z. (1992). Dinosaurian Faunas of China. Beijing: China Ocean Press. p. 187. ISBN 3-540-52084-8.
- ↑ Russell, D. A.; Dong, Z. (1993). "The affinities of a new theropod from the Alxa Desert, Inner Mongolia, People's Republic of China". Canadian Journal of Earth Sciences. 30 (10): 2107−2127. Bibcode:1993CaJES..30.2107R. doi:10.1139/e93-183.
- ↑ Russell, D. A.; Russell, D. E. (1993). "Mammal-dinosaur convergence". National Geographic Research. 9: 70–79. ISSN 8755-724X.
- ↑ Clark, J. M.; Perle, A.; Norell, M. (1994). "The skull of Erlicosaurus andrewsi, a Late Cretaceous Segnosaur (Theropoda, Therizinosauridae) from Mongolia". American Museum Novitates (3115): 1−39. hdl:2246/3712.
- ↑ Nessov, L. A. (1995). Dinosaurs of northern Eurasia: new data about assemblages, ecology, and paleobiogeography (in Russian). Saint Petersburg: Institute of Earth Crust, Saint Petersburg University. p. 49. Translated paper
- ↑ Manning, T. W.; Joysey, K. A.; Cruickshank, A. R. I. (1997). "Observations of microstructures within dinosaur eggs from Henan Province, Peoples' Republic of China". In Wolberg, D. L.; Stump, E.; Rosenberg, R. D. (eds.). Dinofest International: Proceedings of a Symposium Held at Arizona State University. Pennsylvania: Academy of Natural Sciences. pp. 287−290.
- ↑ Dong, Z.; You, H. (1997). "A new segnosaur from Mazhongshan Area, Gansu Province, China". In Dong, Z. M. (ed.). Sino-Japanese Silk Road Dinosaur Expedition. Beijing: China Ocean Press. pp. 90−95.
- ↑ Russell, D. A. (1997). "Therizinosauria". In Currie, P. J.; Padian, K. (eds.). Encyclopedia of Dinosaurs. San Diego: Academic Press. pp. 729−730. ISBN 978-0-12-226810-6.
- ↑ Zhao, X.; Xu, X. (1998). "The oldest coelurosaurian". Nature. 394 (6690): 234–235. Bibcode:1998Natur.394..234Z. doi:10.1038/28300. S2CID 4424059.
- ↑ Xu, X.; Tang, Z.-L.; Wang, X. L. (1999). "A therizinosauroid dinosaur with integumentary structures from China". Nature. 339 (6734): 350−354. Bibcode:1999Natur.399..350X. doi:10.1038/20670. ISSN 1476-4687. S2CID 204993327.
- ↑ Carpener, K. (1999). "The Embryo and Hatching". Eggs, Nests, and Baby Dinosaurs: A Look at Dinosaur Reproduction (Life of the Past). Bloomington: Indiana University Press. p. 209. ISBN 9780253334978.
- ↑ Manning, T. W.; Joysey, K. A.; Cruickshank, A. R. I. (2000). "In ovo tooth replacement in a therizinosauroid dinosaur". In Bravo, A. M.; Reyes, T. (eds.). Extended Abstracts, First International Symposium on Dinosaur Eggs and Babies. Spain: Impremta Provincial de la Diputació Lleida, Isona i Conca Dellà. pp. 129−134.
- ↑ Xu, X.; Zhao, X.; Clark, J. M. (2001). "A new therizinosaur from the Lower Jurassic lower Lufeng Formation of Yunnan, China". Journal of Vertebrate Paleontology. 21 (3): 477−483. doi:10.1671/0272-4634(2001)021[0477:ANTFTL]2.0.CO;2. JSTOR 20061976. S2CID 131298010.
- ↑ Kirkland, J. I.; Wolfe, D. G. (2001). "First definitive therizinosaurid (Dinosauria; Theropoda) from North America". Journal of Vertebrate Paleontology. 21 (3): 410−414. doi:10.1671/0272-4634(2001)021[0410:fdtdtf]2.0.co;2. JSTOR 20061971. S2CID 85705529.
- ↑ Zhang, X.-H.; Xu, X.; Zhao, Z.-J.; Sereno, P. C.; Kuang, X.-W.; Tan, L. (2001). "A long-necked therizinosauroid dinosaur from the Upper Cretaceous Iren Dabasu Formation of Nei Mongol, People's Republic of China" (PDF). Vertebrata PalAsiatica. 39 (4): 282−290.
- ↑ Xu, X.; Zhang, Z.-H.; Sereno, P. C.; Zhao, X.-J.; Kuang, X.-W.; Han, J.; Tan, L. (2002). "A new therizinosauroid (Dinosauria, Theropoda) from the Upper Cretaceous Iren Dabasu Formation of Nei Mongol" (PDF). Vertebrata PalAsiatica. 40: 228−240.
- ↑ Xu, X.; Cheng, Y.; Wang, X.-L.; Chang, C. (2003). "Pygostyle‐like Structure from Beipiaosaurus (Theropoda, Therizinosauroidea) from the Lower Cretaceous Yixian Formation of Liaoning, China". Acta Geologica Sinica. 77 (3): 294–298. doi:10.1111/j.1755-6724.2003.tb00744.x. S2CID 128471217.
- ↑ Kirkland, J. I.; Zanno, L. E.; DeBlieux, D. D.; Sampson, S. D. (2004). "A new, basal-most therizinosauroid (Theropoda: Maniraptora) from Utah demonstrates a pan-Laurasian distribution for Early Cretaceous (Barremian) therizinosauroids". Journal of Vertebrate Paleontology. 24 (supp. 3): 78A. doi:10.1080/02724634.2004.10010643. S2CID 220415208.
- ↑ Smith, D. K.; Kirkland, J. I.; Sanders, R. K.; Zanno, L. E.; DeBlieux, D. D. (2004). "A comparison of North American therizinosaur (Theropoda: Dinosauria) braincases". Journal of Vertebrate Paleontology. 24 (supp. 3): 114A. doi:10.1080/02724634.2004.10010643. S2CID 220415208.
- ↑ Zanno, L. E. (2004). "The pectoral girdle and forelimb of a primitive therizinosauroid (Theropoda: Maniraptora): New information on the phylogenetics and evolution of therizinosaurs". Journal of Vertebrate Paleontology. 24 (supp. 3): 134A. doi:10.1080/02724634.2004.10010643. S2CID 220415208.
- ↑ Zanno, L. E. (2004). The Pectoral Girdle and Forelimb of a Primitive Therizinosauroid (Theropoda, Maniraptora) with Phylogenetic and Functional Implications (PhD diss.). Department of Geology and Geophysics, University of Utah.
- ↑ Kirkland, J. I.; Zanno, L. E.; Sampson, S. D.; Clark, J. M.; DeBlieux, D. D. (2005). "A primitive therizinosauroid dinosaur from the Early Cretaceous of Utah". Nature. 435 (7038): 84–87. Bibcode:2005Natur.435...84K. doi:10.1038/nature03468. PMID 15875020. S2CID 4428196.
- ↑ Zanno, L. E.; Erickson, G. M. (2006). "Ontogeny and life history of Falcarius utahensis, a primitive therizinosauroid from the Early Cretaceous of Utah". Journal of Vertebrate Paleontology. 26 (supp. 3): 143A. doi:10.1080/02724634.2006.10010069. S2CID 220413406.
- ↑ Zanno, L. E. (2006). "The pectoral girdle and forelimb of the primitive therizinosauroid Falcarius Utahensis (Theropoda, Maniraptora): analyzing evolutionary trends within Therizinosauroidea". Journal of Vertebrate Paleontology. 26 (3): 636−650. doi:10.1671/0272-4634(2006)26[636:tpgafo]2.0.co;2. JSTOR 4524610. S2CID 86166623.
- ↑ Burch, S. H. (2006). "The range of motion of the glenohumeral joint of the therizinosaur Neimongosaurus yangi (Dinosauria: Theropoda)". Journal of Vertebrate Paleontology. 26 (supp. 3): 46A. doi:10.1080/02724634.2006.10010069. S2CID 220413406.
- ↑ Sennikov, A. G. (2006). "Читая следы сегнозавров" [Reading segnosaur tracks]. Priroda (in Russian). 5: 58−67.
- ↑ Li, D.; Peng, C.; You, H.; Lamanna, M. C.; Harris, J. D.; Lacovara, K. J.; Zhang, J. (2007). "A Large Therizinosauroid (Dinosauria: Theropoda) from the Early Cretaceous of Northwestern China" (PDF). Acta Geologica Sinica (English Edition). 81 (4): 539–549. doi:10.1111/j.1755-6724.2007.tb00977.x. ISSN 1000-9515. S2CID 130262869.
- ↑ Kundrát, M.; Cruickshank, A. R. I.; Manning, T. W.; Nudds, J. (2007). "Embryos of therizinosauroid theropods from the Upper Cretaceous of China: diagnosis and analysis of ossification patterns". Acta Zoologica. 89 (3): 231−251. doi:10.1111/j.1463-6395.2007.00311.x.
- ↑ Li, D.; You, H.; Zhang, J. (2008). "A new specimen of Suzhousaurus megatherioides (Dinosauria: Therizinosauroidea) from the Early Cretaceous of northwestern China". Canadian Journal of Earth Sciences. 45 (7): 769–779. Bibcode:2008CaJES..45..769L. doi:10.1139/E08-021.
- ↑ Lee, Y.-N.; Barsbold, R.; Currie, P. J. (2008). "A short report of Korea-Mongolia International Dinosaur Project (1st and 2nd year)". Journal of Vertebrate Paleontology. 28 (supp. 003): 104A–105A. doi:10.1080/02724634.2008.10010459.
- ↑ Zanno, L. E.; Gillette, D. D.; Albright, L. B.; Titus, A. L. (2009). "A new North American therizinosaurid and the role of herbivory in predatory dinosaur evolution". Proceedings of the Royal Society B. 276 (1672): 3505−3511. doi:10.1098/rspb.2009.1029. JSTOR 30244145. PMC 2817200. PMID 19605396.
- ↑ Barrett, P. M. (2009). "The affinities of the enigmatic dinosaur Eshanosaurus deguchiianus from the Early Jurassic of Yunnan Province, People's Republic of China". Palaeontology. 52 (4): 681−688. Bibcode:2009Palgy..52..681B. doi:10.1111/j.1475-4983.2009.00887.x.
- ↑ Xu, X.; Zheng, X.; You, H. (2009). "A new feather type in a nonavian theropod and the early evolution of feathers". Proceedings of the National Academy of Sciences. 106 (3): 832−834. doi:10.1073/pnas.0810055106. PMC 2630069. PMID 19139401.
- ↑ Zanno, L. E. (2010). "Osteology of Falcarius utahensis (Dinosauria: Theropoda): characterizing the anatomy of basal therizinosaurs". Zoological Journal of the Linnean Society. 158 (1): 196–230. doi:10.1111/j.1096-3642.2009.00464.x.
- ↑ Zanno, L. E. (2010). "A taxonomic and phylogenetic re-evaluation of Therizinosauria (Dinosauria: Maniraptora)". Journal of Systematic Palaeontology. 8 (4): 503−543. doi:10.1080/14772019.2010.488045. S2CID 53405097.
- ↑ Senter, P.; James, R. H. (2010). "Hip heights of the gigantic theropod dinosaurs Deinocheirus mirificus and Therizinosaurus cheloniformis, and implications for museum mounting and paleoecology" (PDF). Bulletin of Gunma Museum of Natural History (14): 1−10.
- ↑ Smith, D. K.; Zanno, L. E.; Sanders, R. K.; Deblieux, D. D.; Kirkland, J. I. (2011). "New information on the braincase of the North American therizinosaurian (Theropoda, Maniraptora) Falcarius utahensis". Journal of Vertebrate Paleontology. 31 (2): 387−404. Bibcode:2011JVPal..31..387S. doi:10.1080/02724634.2011.549442. JSTOR 25835833. S2CID 73715992.
- ↑ Zanno, L. E.; Makovicky, P. J. (2011). "Herbivorous ecomorphology and specialization patterns in theropod dinosaur evolution". Proceedings of the National Academy of Sciences. 108 (1): 232−237. Bibcode:2011PNAS..108..232Z. doi:10.1073/pnas.1011924108. PMC 3017133. PMID 21173263.
- ↑ Qian, M.-P.; Zhang, Z.-Y.; Jiang, Y.; Jiang, Y.-G.; Zhang, Y.-J.; Chen, R.; Xing, G.-F. (2012). "浙江白垩纪镰刀龙类恐龙" [Cretaceous therizinosaurs in Zhejiang of eastern China]. Journal of Geology (in Chinese). 36 (4): 337−348.
- ↑ Lautenschlager, S.; Emily, J. R.; Perle, A.; Zanno, L. E.; Lawrence, M. W. (2012). "The Endocranial Anatomy of Therizinosauria and Its Implications for Sensory and Cognitive Function". PLOS ONE. 7 (12): e52289. Bibcode:2012PLoSO...752289L. doi:10.1371/journal.pone.0052289. PMC 3526574. PMID 23284972.
- ↑ Senter, P.; Kirkland, J. I.; Deblieux, D. D. (2012). "Martharaptor greenriverensis, a New Theropod Dinosaur from the Lower Cretaceous of Utah". PLOS ONE. 7 (8): e43911. Bibcode:2012PLoSO...743911S. doi:10.1371/journal.pone.0043911. PMC 3430620. PMID 22952806.
- ↑ Fiorillo, A. R.; Adams, T. L. (2012). "A Therizinosaur Track from the Lower Cantwell Formation (upper Cretaceous) of Denali National Park, Alaska". PALAIOS. 27 (6): 395−400. Bibcode:2012Palai..27..395F. doi:10.2110/palo.2011.p11-083r. S2CID 129517554.
- ↑ Lautenschlager, S. (2013). "Cranial myology and bite force performance of Erlikosaurus andrewsi : a novel approach for digital muscle reconstructions". Journal of Anatomy. 222 (2): 260−272. doi:10.1111/joa.12000. PMC 3632231. PMID 23061752.
- ↑ Lautenschlager, S.; Witmer, L. M.; Perle, A.; Rayfield, E. J. (2013). "Edentulism, beaks, and biomechanical innovations in the evolution of theropod dinosaurs". PNAS. 110 (51): 20657−20662. Bibcode:2013PNAS..11020657L. doi:10.1073/pnas.1310711110. JSTOR 23761610. PMC 3870693. PMID 24297877.
- ↑ Pu, H.; Kobayashi, Y.; Lü, J.; Xu, L.; Wu, Y.; Chang, H.; Zhang, J.; Jia, S. (2013). "An Unusual Basal Therizinosaur Dinosaur with an Ornithischian Dental Arrangement from Northeastern China". PLOS ONE. 8 (5): e63423. Bibcode:2013PLoSO...863423P. doi:10.1371/journal.pone.0063423. PMC 3667168. PMID 23734177.
- ↑ Kobayashi, Y.; Lee, Y.; Barsbold, R.; Zelenitsky, D.; Tanaka, K. (2013). "First record of a dinosaur nesting colony from Mongolia reveals nesting behavior of therizinosauroids". In Maxwell, E.; Miller-Camp, J.; Anemone, R. (eds.). 73rd Annual Meeting of Society of Vertebrate Paleontology. Los Angeles. p. 155.
{{cite book}}: CS1 maint: location missing publisher (link) - ↑ Lautenschlager, S. (2014). "Morphological and functional diversity in therizinosaur claws and the implications for theropod claw evolution". Proceedings of the Royal Society B. 28 (1785): 20140497. doi:10.1098/rspb.2014.0497. PMC 4024305. PMID 24807260.
- ↑ Li, Q.; Clarke, J. A.; Gao, K.-Q.; Zhou, C.-F.; Meng, Q.; Li, D.; D’Alba, L.; Shawkey, M. D. (2014). "Melanosome evolution indicates a key physiological shift within feathered dinosaurs". Nature. 507 (7492): 350−353. Bibcode:2014Natur.507..350L. doi:10.1038/nature12973. PMID 24522537. S2CID 4395833.
- ↑ Hedrick, B. P.; Zanno, L. E.; Wolfe, D. G.; Dodson, P. (2015). "The Slothful Claw: Osteology and Taphonomy of Nothronychus mckinleyi and N. graffami (Dinosauria: Theropoda) and Anatomical Considerations for Derived Therizinosaurids". PLOS ONE. 10 (6): e0129449. Bibcode:2015PLoSO..1029449H. doi:10.1371/journal.pone.0129449. PMC 4465624. PMID 26061728.
- ↑ Smith, D. K. (2015). "Craniocervical Myology and Functional Morphology of the Small-Headed Therizinosaurian Theropods Falcarius utahensis and Nothronychus mckinleyi". PLOS ONE. 10 (2): e0117281. Bibcode:2015PLoSO..1017281S. doi:10.1371/journal.pone.0117281. PMC 4338109. PMID 25706764.
- ↑ Kobayashi, Y.; Chinzorig, T.; Tsogtbaatar, K.; Barsbold, R. (2015). "A new therizinosaur with functionally didactyl hands from the Bayanshiree Formation (Cenomanian-Turonian), Omnogovi Province, southeastern Mongolia" (PDF). Journal of Vertebrate Paleontology. Program and Abstracts. Photographs
- ↑ Gierliński, G. D. (2015). "New Dinosaur Footprints from the Upper Cretaceous of Poland in the Light of Paleogeographic Context". Ichnos. 22 (3–4): 220−226. Bibcode:2015Ichno..22..220G. doi:10.1080/10420940.2015.1063489. S2CID 128598690.
- ↑ Zanno, L. E.; Tsogtbaatar, K.; Chinzorig, T.; Gates, T. A. (2016). "Specializations of the mandibular anatomy and dentition of Segnosaurus galbinensis (Theropoda: Therizinosauria)". PeerJ. 4: e1885. doi:10.7717/peerj.1885. PMC 4824891. PMID 27069815.
- ↑ Sues, H.-D.; Averianov, A. (2016). "Therizinosauroidea (Dinosauria: Theropoda) from the Upper Cretaceous of Uzbekistan". Cretaceous Research. 59: 155−178. Bibcode:2016CrRes..59..155S. doi:10.1016/j.cretres.2015.11.003.
- ↑ Botelho, J. H.; Smith-Paredes, D.; Soto-Acuña, S.; Núñez-León, D.; Palma, V.; Vargas, A. O. (2016). "Greater Growth of Proximal Metatarsals in Bird Embryos and the Evolution of Hallux Position in the Grasping Foot". Journal of Experimental Zoology Part B: Molecular and Developmental Evolution. 328 (1–2): 106−118. doi:10.1002/jez.b.22697. hdl:10533/232005. ISSN 1552-5015. PMID 27649924. S2CID 4952929.
- ↑ Lautenschlager, S. (2017). "Functional niche partitioning in Therizinosauria provides new insights into the evolution of theropod herbivory" (PDF). Palaeontology. 60 (3): 375−387. Bibcode:2017Palgy..60..375L. doi:10.1111/pala.12289. S2CID 90965431.
- ↑ Masrour, M.; Lkebir, N.; Pérez-Lorente, F. (2017). "Anza palaeoichnological site. Late Cretaceous. Morocco. Part II. Problems of large dinosaur trackways and the first African Macropodosaurus trackway". Journal of African Earth Sciences. 134: 776−793. Bibcode:2017JAfES.134..776M. doi:10.1016/j.jafrearsci.2017.04.019. ISSN 1464-343X.
- ↑ Button, K.; You, H.; Kirkland, J. I.; Zanno, L. E. (2017). "Incremental growth of therizinosaurian dental tissues: implications for dietary transitions in Theropoda". PeerJ. 5: e4129. doi:10.7717/peerj.4129. PMC 5729821. PMID 29250467.
- ↑ Smith, D. K.; Sanders, R. K.; Wolfe, D. G. (2018). "A re-evaluation of the basicranial soft tissues and pneumaticity of the therizinosaurian Nothronychus mckinleyi (Theropoda; Maniraptora)". PLOS ONE. 13 (7): e0198155. Bibcode:2018PLoSO..1398155S. doi:10.1371/journal.pone.0198155. PMC 6067709. PMID 30063717.
- ↑ Fiorillo, A. R.; McCarthy, P. J.; Kobayashi, Y.; Tomsich, C. S.; Tykoski, R. S.; Lee, Y.-N.; Tanaka, T.; Noto, C. R. (2018). "An unusual association of hadrosaur and therizinosaur tracks within Late Cretaceous rocks of Denali National Park, Alaska". Scientific Reports. 8 (11706): 11706. Bibcode:2018NatSR...811706F. doi:10.1038/s41598-018-30110-8. PMC 6076232. PMID 30076347.
- ↑ McNamara, M. E.; Zhang, F.; Kearns, S. L.; Orr, P. J.; Toulouse, A.; Foley, T.; Hone, D. W. E; Rogers, C. S.; Benton, M. J.; Johnson, D.; Xu, X.; Zhou, Z. (2018). "Fossilized skin reveals coevolution with feathers and metabolism in feathered dinosaurs and early birds". Nature Communications. 9 (2072): 2072. Bibcode:2018NatCo...9.2072M. doi:10.1038/s41467-018-04443-x. ISSN 2041-1723. PMC 5970262. PMID 29802246.
- ↑ Macaluso, L.; Tschopp, E.; Mannion, P. (2018). "Evolutionary changes in pubic orientation in dinosaurs are more strongly correlated with the ventilation system than with herbivory". Palaeontology. 61 (5): 703−719. Bibcode:2018Palgy..61..703M. doi:10.1111/pala.12362. S2CID 133643430.
- ↑ Hartman, S.; Mortimer, M.; Wahl, W. R.; Lomax, D. R.; Lippincott, J.; Lovelace, D. M. (2019). "A new paravian dinosaur from the Late Jurassic of North America supports a late acquisition of avian flight". PeerJ. 7: e7247. doi:10.7717/peerj.7247. PMC 6626525. PMID 31333906.
- ↑ Xi Yao; Chun-Chi Liao; Corwin Sullivan; Xing Xu (2019). "A new transitional therizinosaurian theropod from the Early Cretaceous Jehol Biota of China". Scientific Reports. 9 (1): Article number 5026. Bibcode:2019NatSR...9.5026Y. doi:10.1038/s41598-019-41560-z. PMC 6430829. PMID 30903000.
- ↑ Liao, C.-C.; Xu, X. (2019). "Cranial osteology of Beipiaosaurus inexpectus (Theropoda: Therizinosauria)". Vertebrata PalAsiatica. 57 (2): 117–132. doi:10.19615/j.cnki.1000-3118.190115.
- ↑ Nabavizadeh, A. (2019). "Cranial musculature in herbivorous dinosaurs: a survey of reconstructed anatomical diversity and feeding mechanisms". The Anatomical Record. 303 (4): 1104–1145. doi:10.1002/ar.24283. PMID 31675182. S2CID 207815224.
- ↑ Button, D. J.; Zanno, L. E. (2019). "Repeated evolution of divergent modes of herbivory in non-avian dinosaurs". Current Biology. 30 (1): 158−168.e4. doi:10.1016/j.cub.2019.10.050. PMID 31813611. S2CID 208652510.
- ↑ Tanaka, K.; Kobayashi, Y.; Zelenitsky, D. K.; Therrien, F.; Lee, Y.-N.; Barsbold, R.; Kubota, K.; Lee, H.-J.; Tsogtbaatar, C.; Idersaikhan, D. (2019). "Exceptional preservation of a Late Cretaceous dinosaur nesting site from Mongolia reveals colonial nesting behavior in a non-avian theropod". Geology. 47 (9): 843−847. Bibcode:2019Geo....47..843T. doi:10.1130/G46328.1. S2CID 198412503.
- ↑ Smith, D. K.; Sanders, R. K.; Wolfe, D. G. (2020). "Vertebral pneumaticity of the North American therizinosaur Nothronychus". Journal of Anatomy. 238 (3): 598−614. doi:10.1111/joa.13327. PMC 7855063. PMID 33044012.
- ↑ Smith, D. K. (2021). "Hind limb muscle reconstruction in the incipiently opisthopubic large therizinosaur Nothronychus (Theropoda; Maniraptora)". Journal of Anatomy. 238 (6): 1404–1424. doi:10.1111/joa.13382. PMC 8128771. PMID 33417263.
- ↑ Smith, D. K. (2021). "Forelimb musculature and function in the therizinosaur Nothronychus (Maniraptora, Theropoda)". Journal of Anatomy. 239 (2): 307–335. doi:10.1111/joa.13418. PMC 8273597. PMID 33665832.
- ↑ Liao, C.-C.; Zanno, L. E.; Wang, S.; Xu, X. (2021). "Postcranial osteology of Beipiaosaurus inexpectus (Theropoda: Therizinosauria)". PLOS ONE. 16 (9): e0257913. Bibcode:2021PLoSO..1657913L. doi:10.1371/journal.pone.0257913. PMC 8483305. PMID 34591927.
- ↑ Hattori, S.; Kawabe, S.; Imai, T.; Shibata, M.; Miyata, K.; Xu, X.; Azuma, Y. (2021). "Osteology of Fukuivenator paradoxus: a bizarre maniraptoran theropod from the Early Cretaceous of Fukui, Japan" (PDF). Memoir of the Fukui Prefectural Dinosaur Museum. 20: 1–82.
- ↑ Kundrát, M.; Cruickshank, A. R. (2023). "New information on multispherulitic dinosaur eggs: Faveoloolithidae and Dendroolithidae". Historical Biology. 34 (6): 1072−1084. doi:10.1080/08912963.2021.1961764. S2CID 238687752.
- ↑ Sennikov, A. G. (2021). "The Plantigrade Segnosaurians: Sloth Dinosaurs or Bear Dinosaurs?". Paleontological Journal. 55 (7): 1158–1185. doi:10.1134/S0031030121100087. S2CID 245539994.
- ↑ Kobayashi, Y.; Takasaki, R.; Fiorillo, A. R.; Chinzorig, T.; Hikida, Y. (2022). "New therizinosaurid dinosaur from the marine Osoushinai Formation (Upper Cretaceous, Japan) provides insight for function and evolution of therizinosaur claws". Scientific Reports. 12 (7207): 7207. Bibcode:2022NatSR..12.7207K. doi:10.1038/s41598-022-11063-5. PMC 9065154. PMID 35504901.
- ↑ Qin, Z.; Liao, C.-C.; Benton, M. J.; Rayfield, E. J. (2023). "Functional space analyses reveal the function and evolution of the most bizarre theropod manual unguals". Communications Biology. 6 (181): e12640. doi:10.1038/s42003-023-04552-4. PMC 9935540. PMID 36797463.
- ↑ Smith, D. K.; Gillette, D. D. (2023). "Reconstruction of soft noncontractile tissue in the derived therizinosaur Nothronychus: The interplay of soft tissue and stress on hindlimb ossification and posture". Journal of Morphology. 284 (5): e21579. doi:10.1002/jmor.21579. PMID 36929022. S2CID 257583565.
- ↑ Smith, D. K. (2023). "Hindlimb Musculature of the Lower Cretaceous (Barremian) Therizinosaur Falcarius utahensis (Maniraptora, Theropoda) with Implications for Evolution, Stance, and Stride". Cretaceous Research. 149: 105557. Bibcode:2023CrRes.14905557S. doi:10.1016/j.cretres.2023.105557. S2CID 258177949.
- ↑ Wills, S.; Underwood, C. J.; Barrett, P. M. (2023). "Machine learning confirms new records of maniraptoran theropods in Middle Jurassic UK microvertebrate faunas". Papers in Palaeontology. 9 (2): e1487. doi:10.1002/spp2.1487.