| ZC3H12B | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Identifiers | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Aliases | ZC3H12B, CXorf32, MCPIP2, zinc finger CCCH-type containing 12B | ||||||||||||||||||||||||||||||||||||||||||||||||||
| External IDs | OMIM: 300889 MGI: 2442133 HomoloGene: 19395 GeneCards: ZC3H12B | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Wikidata | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
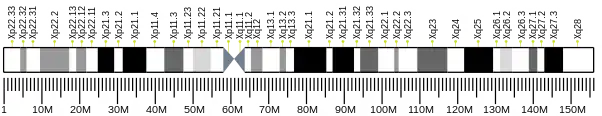

ZC3H12B, also known as CXorf32 or MCPIP2, is a protein encoded by gene ZC3H12B located on chromosome Xq12 in humans.
Gene
The ZC3H12B gene is composed of 19,709 base pairs (bp) and contains 5 exons. It is located on the X chromosome at q12 on the plus strand.
 ZC3H12B Locus.
ZC3H12B Locus.
ZC3H12B contains a ribonuclease domain, as well as a CCCH-type zinc finger domain. Ribonucleases (RNases) degrade RNA and are involved in the RNA maturation process. They are also a line of defense against viral RNA (D'Alessio and Riordan 1997). CCCH-type zinc fingers are associated with mRNA destabilization. CCCH-type zinc fingers have been shown to turn over mRNA without the removal of the PolyA tail (Lai and Blackshear 2001). ZC3H12B and its paralogs ZC3H12A, ZC3H12C and ZC3H12D all contain CCCH-type zinc finger domains, which have been associated with cell cycle and growth phase transitions in eukaryotes (InterPro).
Promoter
Genomatix ElDorado program predicted a 601 bp promoter upstream of the ZC3H12B gene with multiple transcription factor binding sites including nuclear factor of activated T-cells and ribonucleoprotein associated zinc finger protein MOK-2 (also known as ZNF239).
 Transcription factor binding site predictions in the promoter of ZC3H12B.
Transcription factor binding site predictions in the promoter of ZC3H12B.
mRNA
ZC3H12B contains 7273 bp mRNA. There is only one predicted transcript by Aceview. No folding patterns have been predicted (Mfold). There are for introns excised from ZC3H12B.
 Aceview single-predicted mRNA variant in ZC3H12B.
Aceview single-predicted mRNA variant in ZC3H12B.
Protein
ZC3H12B is a probable ribonuclease containing CCCH-type zinc finger domain and ribonuclease domains. The 836 amino acid protein has a predicted molecular weight of 94.2 kdal. It does not contain a signal peptide or a transmembrane region. PSORTII predicted 65.2% probability of nuclear location. CCCH-type zinc fingers and ribonucleases are presumably located in the nucleus for RNA cleaving and specifically, RNA hairpin cleaving (Boysen and Hearn 2008).
Structural characteristics
The protein secondary structure is a mixture of alpha helices and beta strands. The two domains identified so far are the ribonuclease and CCCH-type zinc finger domains.
Shown below is a conserved domain of ZC3H12B paralog, Mcpip1 (or ZC3H12A). In a BLAST structure comparison, there was an 82% identity match with 24% query coverage, with a predicted e-value of 2e-118. 82% identity match is enough to make comparisons of ZC3H12B and Mcpip1 (ZC3H12A) zinc finger conserved domain, which are both predicted to be composed of beta strands and alpha helices.
 Cn3D rendering of Mcpip1 conserved domain with zinc-finger motif.
Cn3D rendering of Mcpip1 conserved domain with zinc-finger motif.
Post-translational modification
Phobius program predicted non-cytoplasmic protein location. NetPhos 2.0. predicted 63 phosphorylation sites in ZC3H12B, which are marked on the conceptual translation. YinOYang1.2. predicted three 0-Beta-GlcNAc attachment sites, which are competing with phosphorylation sites. 0-Beta-GlcNAc is presumably the only type of glycosylation occurring in the nucleus and/or cytoplasm of cells. There is a notable link between antigen activation by lymphocytes and dynamic 0-B-Glycosylation in nuclear proteins (Hart and Akimoto). NetNGlyc predicted glycosylation sites; however, these sites were excluded because the protein is likely nuclear and would not undergo this form of glycosylation. There were no predicted acetylation sites at the N-terminus of the protein. This is unusual because approximately 85% of human proteins are acetylated at the N terminus for synthesis, stabilization and localization of proteins (Van Damm et al.). No positive, negative or mixed charge clusters present. No hydrophobic segments detected (SAPS SDSC Biology Workbench). MitoProtII did not detect any mitochondria export signals. These post-translational tests suggest the protein is located in the nucleus and undergoes dynamic phosphorylation and 0-Beta-GlcNAc modifications.
Evolution
Select domains of ZC3H12B are conserved in most vertebrates, arthropods and annelids. There are not conserved domains in domains bacteria or archaea. There were not significantly conserved domains in yeasts, plants or protists.
Paralogs
There are three paralogs of ZC3H12B which are in the same CCCH-type zinc finger family, all which maintain greater than 50% identity to ZC3H12B based on BLAST analysis (NCBI).
| Name | Species | NCBI accession number | Length (AA) | Protein identity |
|---|---|---|---|---|
| ZC3H12B | Homo sapiens | NM_001010888.3 | 836aa | 100% |
| ZC3H12A | Homo sapiens | NM_025079.2 | 599aa | 68% |
| ZC3H12C | Homo sapiens | NM_033390.1 | 883aa | 53% |
| ZC3H12D | Homo sapiens | NM_207360.2 | 527aa | 61% |
Orthologs
ZC3H12B is conserved in mammals, birds, insects and nematodes (BLAST). See the table below for the summary of orthologs of ZC3H12B in humans.
| Species | Species common name | Divergence (MYA) | NCBI accession number (protein) | Length (amino acids) | Protein identity | Similarity | |
|---|---|---|---|---|---|---|---|
| Homo sapiens | Human | n/a | NP_001010888.3 | 836aa | 100% | 100% | |
| Pan paniscus | Chimpanzee | 6.3 | XP_003816967.1 | 836aa | 99% | 99% | |
| Pongo abelii | Orangutan | 15.7 | XP_002831786.1 | 836aa | 99% | 99% | |
| Macaca mulatta | Rhesus monkey | 29 | XP_002806307.1 | 836aa | 99% | 99% | |
| Callithrix jacchus | Marmoset | 42.6 | XP_002762992.2 | 836aa | 98% | 98% | |
| Mus musculus | Mouse | 92.3 | NP_001030079.2 | 835aa | 91% | 94% | |
| Sus scrofa | Pig | 94.2 | XP_003360389.1 | 836aa | 93% | 96% | |
| Gallus gallus | Chicken | 296 | XP_003641177.1 | 837aa | 77% | 85% | |
| Chrysemys picta bellii | Painted turtle | 296 | XP_005279572.1 | 838aa | 78% | 86% | |
| Oryzias latipes | Medaka | 400.1 | XP_004076599.1 | 845aa | 67% | 77% | |
| Gadus morhua | Atlantic cod | 400.1 | AFK76491.1 | 842aa | 29% | 44% | |
| Danio rerio | Zebrafish | 400.1 | XP_001342172.3 | 982aa | 68% | 77% | |
| Petromyzon marinus | Lamprey | 535.7 | ABO21295.1 | 222aa | 44% | 58% | |
| Branchiostoma floridae | Lancelet | 713.2 | XP_002598834.1 | 492aa | 66% | 79% | |
| Ciona intestinalis | Vase tunicate | 722.5 | XP_002125834.1 | 863aa | 54% | 66% | |
| Strongylocentrotus purpuratus | Purple sea urchin | 742.9 | XP_787030.3 | 974aa | 58% | 72% | |
| Aplysiomorpha californica | Sea hare | 782.7 | XP_005113312.1 | 1269aa | 51% | 69% | |
| Drosophila grimshawi | Hawaii fruit fly | 782.7 | XP_001994140.1 | 548aa | 51% | 69% | |
| Anopheles gambiae | Mosquito | 782.7 | XP_321880 | 637aa | 59% | 75% | |
| Apis mellifera | Honey bee | 782.7 | XP_397264 | 652aa | 58% | 73% | |
| Caenorhabditis elegans | Round worm (nematode) | 937.5 | NP_491985.4 | 634aa | 46% | 64% |
Expression and function
Microarrays in normal tissue expression profiling showed increased expression of the gene in the pancreas, prostate, brain, spinal cord and thymus (GEO). Unlike its paralogs, it is not expressed in macrophage-activated tissues, which indicates the paralogous relationship to the inflammatory response (Liang et al. 2008). ZC3H12B is expressed transiently in brain, thymus and testis tissues (EST).
Interaction
Predicted interactions by Ingenuity Systems showed no drug targeting molecules in pathway and no known drug targets. The listed top functions and diseases were cancer, organismal injury and abnormalities, reproductive system disease. Several miRNA interactions were predicted. The predicted miRNA targets have yet to be matched to the ZC3H12B sequence and it is unclear whether there is an interaction between the two. Tests such as Forster Resonance Energy Transfer (FRET), co-immunoprecipitation, two-hybrid screening, hydropathic complementarity, cluster-microarray and ChiP could be used in the future to test for new protein/chromatin interactions with ZC3H12B.
Clinical significance
Deletions of the Xq12 locus has resulted in several disorders such as androgen insensitivity, susceptibility to prostate cancer, spinal and bulbar muscular atrophy of Kennedy and mental retardation; however, no link has been found between these diseases and ZC3H12B (NCBI).
References
- 1 2 3 GRCh38: Ensembl release 89: ENSG00000102053 - Ensembl, May 2017
- 1 2 3 GRCm38: Ensembl release 89: ENSMUSG00000035045 - Ensembl, May 2017
- ↑ "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- ↑ "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
Further reading
- Liang J, Wang J, Azfer A, Song W, Tromp G, Kolattukudy PE, Fu M (March 2008). "A novel CCCH-zinc finger protein family regulates proinflammatory activation of macrophages". The Journal of Biological Chemistry. 283 (10): 6337–46. doi:10.1074/jbc.m707861200. PMID 18178554.
- Van Damme P, Hole K, Pimenta-Marques A, Helsens K, Vandekerckhove J, Martinho RG, Gevaert K, Arnesen T (July 2011). "NatF contributes to an evolutionary shift in protein N-terminal acetylation and is important for normal chromosome segregation". PLOS Genetics. 7 (7): e1002169. doi:10.1371/journal.pgen.1002169. PMC 3131286. PMID 21750686.