
Cysteinyl leukotriene receptor 1, also termed CYSLTR1, is a receptor for cysteinyl leukotrienes (LT) (see cysteinyl leukotrienes). CYSLTR1, by binding these cysteinyl LTs (CysLTs; viz, LTC4, LTD4, and to a much lesser extent, LTE4) contributes to mediating various allergic and hypersensitivity reactions in humans as well as models of the reactions in other animals.
Gene
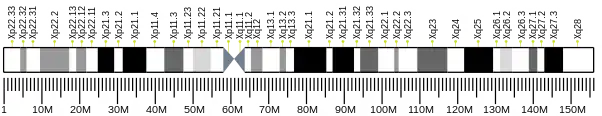
The human CysLTR1 gene maps to the X chromosome at position Xq13-Xq21, contains three exons with the entire open reading frame located in exon 3, and codes for a protein composed of 337 amino acids. The CYSLTR1 gene promoter region is distanced from 665 to 30 bp upstream of its transcription start site.[5][6][7]
Expression
CYSLTR1 mRNA is expressed in lung smooth muscle, lung macrophages, monocytes, eosinophils, basophils, neutrophils, platelets, T cells, B lymphocytes, pluripotent hematopoietic stem cells (CD34+), mast cells, pancreas, small intestine, prostate, interstitial cells of the nasal mucosa, airway smooth muscle cells, bronchial fibroblasts and vascular endothelial cells.[5][6][8]
Function
CysLTR1 is a G protein–coupled receptor that links to and when bound to its CysLT ligands activates the Gq alpha subunit and/or Ga subunit of its coupled G protein, depending on the cell type. Acting through these G proteins and their subunits, ligand-bound CysLTR1 activates a series of pathways that lead to cell function; the order of potency of the cysLTs in stimulating CysLTR1 is LTD4>LTC4>LTE4[7] with LTE4 probably lacking sufficient potency to have much activity that operates through CysLTR1 in vivo.[6]
CysLTR1 activation by LTC4 and/or LTD4 in animal models and humans causes: airway bronchoconstriction and hyper-responsiveness to bronchoconstriction agents such as histamine; increased vascular permeability, edema, influx of eosinophils and neutrophils, smooth muscle proliferation, collagen deposition, and fibrosis in various tissue sites; and mucin secretion by goblet cells, goblet cell metaplasia, and epithelial cell hypertrophy in the membranes of the respiratory system.[7] Animal model and human tissue (preclinical studies) implicate CysLTR1 antagonists as having protective/reparative effects in models of brain injury (trauma-, ischemia-, and cold-induced), multiple sclerosis, auto-immune encephalomyelitis, Alzheimer's disease, and Parkinson's disease.[9] CysLTR1 activation is also associated in animal models with decreasing the blood–brain barrier (i.e. increasing the permeability of brain capillaries to elements of the blood's soluble elements) as well as promoting the movement of leukocytes for the blood to brain tissues; these effects may increase the development and frequency of epileptic seizure as well as the entry of leucocyte-borne viruses such as HIV-1 into brain tissue.[9]
Increased expression of CysLTR1 has been observed in transitional cell carcinoma of the urinary bladder, neuroblastoma and other brain cancers, prostate cancer, breast cancer, and colorectal cancer (CRC); indeed, CysLTR1 tumor expression is associated with poor survival prognoses in breast cancer and CRC patients, and drug inhibitors of CysLTR1 block the in vivo and in vivo (animal model) growth of CRC cells and tumors, respectively. The pro-cancer effects of CysLTR1 in CRC appear due to its ability to up-regulate pathways that increase in CRC cell proliferation and survival.[10][11]
Other cysLT receptors include cysteinyl leukotriene receptor 2 (i.e. CysLTR2) and GPR99 (also termed the oxoglutarate receptor and, sometimes, CysLTR3).[6] The order of potency of the cysLTs in stimulating CysLTR2 is LTD4=LTC4>LTE4[7] with LTE4 probably lacking sufficient potency to have much activity that operates through CysLTR2 in vivo.[6] GPR99 appears to be an important receptor for CysLTs, particularly for LTE4. The CysLTs show relative potencies of LTE4>LTC4>LTD4 in stimulating GPR99-bearing cells with GPR99-deficient mice exhibiting a dose-dependent loss of vascular permeability responses in skin to LTE4 but not to LTC4 or LTD4. This and other data suggest that GPR99 is an important receptor for the in vivo actions of LTE4 but not LTD4 or LTC4[6][12][13]
The GPR17 receptor, also termed the uracil nucleotide/cysteinyl leukotriene receptor, was initially defined as a receptor for LTC4, LTD4, and uracil nucleotides. However, more recent studies from different laboratories could not confirm these results; they found that GPR17-bearing cells did not respond to these CysLTs or nucleotides but did find that cells expressing both CysLTR1 and GPR17 receptors exhibited a marked reduction in binding LTC4 and that mice lacking GPR17 were hyper-responsive to igE-induced passive cutaneous anaphylaxis. GPR17 therefore appears to inhibit CysLTR1, at least in these model systems.[14] In striking contrast to these studies, studies concentration on neural tissues continue to find that Oligodendrocyte progenitor cells express GPR17 and respond through this receptor to LTC4, LTD4, and certain purines (see GPR17#Function).
The Purinergic receptor, P2Y12, while not directly binding or responding to CysLTs, appears to be activated as a consequence of activating CysLT1: blockage of P2Y12 activation either by receptor depletion or pharmacological methods inhibits many of the CysLTR1-dependent actions of CysLTs in various cell types in vitro as well as in an animal model of allergic disease.[8][15][16][9]
Ligands
The major CysLTs viz., LTC4, LTD4, and LTE4, are metabolites of arachidonic acid made by the 5-lipoxygenase enzyme, ALOX5, mainly by cells involved in regulating inflammation, allergy, and other immune responses such as neutrophils, eosinophils, basophils, monocytes, macrophages, mast cells, dendritic cells, and B-lymphocytes. ALOX5 metabolizes arachidonic acid to the 5,6-epoxide precursor, LTA4, which is then acted on by LTC4 synthase which attaches the γ-glutamyl-cysteinyl-glycine tripeptide (i.e. glutathione) to carbon 6 of the intermediate thereby forming LTC4 synthase. LTC4 then exits its cells of origin through the MRP1 transporter (ABCC1) and is rapidly converted to LTD4 and then to LTE4) by cell surface-attached gamma-glutamyltransferase and dipeptidase peptidase enzymes by the sequential removal of the γ-glutamyl and then glycine residues.[7][17][18]
Gene polymorphism
927T/C (nucleotide thymine replaces cytosine at position 97 of the CysLTR1 gene) gene polymorphism in the coding region of CysLTR1 has been shown to be predictive of the severity of atopy (i.e. a predisposition toward developing certain allergic hypersensitivity reactions), but not associated with asthma, in a population of 341 Caucasians in afflicted sib-pair families from the Southampton area in the United Kingdom. This atopy severity was most apparent in female siblings but the incidence of this polymorphism is extremely low and the functionality of the 927T/C gene and its product protein are as yet unknown.[19][20]
The population of the small remote far South Atlantic Ocean island of Tristan da Cunha (266 permanent, genetically isolated residents) suffers a high prevalence of atopy and asthma. The CysLTR1 gene product variant, 300G/S (i.e. amino acid glycine replaces serine at the 300 position of the CysLTR1 protein), has been shown to be significantly associated with atopy in this population. The CysLTR1 300S variant exhibited significant increased sensitivity to LTD4 and LTC4 suggesting that this hypersensitivity underlies its association with atopy.[21][22]
Clinical significance
In spite of the other receptors cited as being responsive to CysLTs, CysLTR1 appears to be critical in mediating many of the pathological responses to CysLTs in humans. Montelukast, Zafirlukast, and Pranlukast are selective receptor antagonists for the CysLTR1 but not CysLTR2. These drugs are in use and/or shown to be effective as prophylaxis and chronic treatments for allergic and non-allergic diseases such as: allergen-induced asthma and rhinitis; aspirin-exacerbated respiratory disease; exercise- and cold-air induced asthma (see Exercise-induced bronchoconstriction); and childhood sleep apnea due to adenotonsillar hypertrophy (see Acquired non-inflammatory myopathy#Diet and Trauma Induced Myopathy).[17][18][23][24] However, responses to these lukast drugs vary greatly with the drugs showing fairly high rates of poor responses and ~20% of patients reporting no change in symptoms after treatment with these agents.[13][25][26] It seems possible that the responses of CysLTR2, GPR99, or other receptors to CysLT's may be contributing to these diseases.[8][15]
See also
References
- 1 2 3 GRCh38: Ensembl release 89: ENSG00000173198 - Ensembl, May 2017
- 1 2 3 GRCm38: Ensembl release 89: ENSMUSG00000052821 - Ensembl, May 2017
- ↑ "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- ↑ "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- 1 2 Zhang J, Migita O, Koga M, Shibasaki M, Arinami T, Noguchi E (June 2006). "Determination of structure and transcriptional regulation of CYSLTR1 and an association study with asthma and rhinitis". Pediatric Allergy and Immunology. 17 (4): 242–9. doi:10.1111/j.1399-3038.2005.00347.x. PMID 16771777. S2CID 23133928.
- 1 2 3 4 5 6 Singh RK, Tandon R, Dastidar SG, Ray A (November 2013). "A review on leukotrienes and their receptors with reference to asthma". The Journal of Asthma. 50 (9): 922–31. doi:10.3109/02770903.2013.823447. PMID 23859232. S2CID 11433313.
- 1 2 3 4 5 Liu M, Yokomizo T (2015). "The role of leukotrienes in allergic diseases". Allergology International. 64 (1): 17–26. doi:10.1016/j.alit.2014.09.001. PMID 25572555.
- 1 2 3 Cattaneo M (2015). "P2Y12 receptors: structure and function". Journal of Thrombosis and Haemostasis. 13 (Suppl 1): S10–6. doi:10.1111/jth.12952. PMID 26149010. S2CID 206159450.
- 1 2 3 Ghosh A, Chen F, Thakur A, Hong H (2016). "Cysteinyl Leukotrienes and Their Receptors: Emerging Therapeutic Targets in Central Nervous System Disorders". CNS Neuroscience & Therapeutics. 22 (12): 943–951. doi:10.1111/cns.12596. PMC 6492851. PMID 27542570.
- ↑ Singh RK, Gupta S, Dastidar S, Ray A (2010). "Cysteinyl leukotrienes and their receptors: molecular and functional characteristics". Pharmacology. 85 (6): 336–49. doi:10.1159/000312669. PMID 20516735.
- ↑ Savari S, Vinnakota K, Zhang Y, Sjölander A (2014). "Cysteinyl leukotrienes and their receptors: bridging inflammation and colorectal cancer". World Journal of Gastroenterology. 20 (4): 968–77. doi:10.3748/wjg.v20.i4.968. PMC 3921548. PMID 24574769.
- ↑ Bankova LG, Lai J, Yoshimoto E, Boyce JA, Austen KF, Kanaoka Y, Barrett NA (May 2016). "Leukotriene E4 elicits respiratory epithelial cell mucin release through the G-protein-coupled receptor, GPR99". Proceedings of the National Academy of Sciences of the United States of America. 113 (22): 6242–7. Bibcode:2016PNAS..113.6242B. doi:10.1073/pnas.1605957113. PMC 4896673. PMID 27185938.
- 1 2 Kanaoka Y, Maekawa A, Austen KF (April 2013). "Identification of GPR99 protein as a potential third cysteinyl leukotriene receptor with a preference for leukotriene E4 ligand". The Journal of Biological Chemistry. 288 (16): 10967–72. doi:10.1074/jbc.C113.453704. PMC 3630866. PMID 23504326.
- ↑ Kanaoka Y, Boyce JA (2014). "Cysteinyl leukotrienes and their receptors; emerging concepts". Allergy, Asthma & Immunology Research. 6 (4): 288–95. doi:10.4168/aair.2014.6.4.288. PMC 4077954. PMID 24991451.
- 1 2 Austen KF, Maekawa A, Kanaoka Y, Boyce JA (2009). "The leukotriene E4 puzzle: finding the missing pieces and revealing the pathobiologic implications". The Journal of Allergy and Clinical Immunology. 124 (3): 406–14, quiz 415–6. doi:10.1016/j.jaci.2009.05.046. PMC 2739263. PMID 19647860.
- ↑ Baptista-dos-Reis R, Muniz VS, Neves JS (2015). "Multifaceted roles of cysteinyl leukotrienes in eliciting eosinophil granule protein secretion". BioMed Research International. 2015: 1–7. doi:10.1155/2015/848762. PMC 4383494. PMID 25866815.
- 1 2 Haeggström JZ, Funk CD (October 2011). "Lipoxygenase and leukotriene pathways: biochemistry, biology, and roles in disease". Chemical Reviews. 111 (10): 5866–98. doi:10.1021/cr200246d. PMID 21936577.
- 1 2 Anwar Y, Sabir JS, Qureshi MI, Saini KS (April 2014). "5-lipoxygenase: a promising drug target against inflammatory diseases-biochemical and pharmacological regulation". Current Drug Targets. 15 (4): 410–22. doi:10.2174/1389450114666131209110745. PMID 24313690.
- ↑ Allen BW (1980). "Inhibitory effect of rubber on the growth of M. tuberculosis". Tubercle. 61 (2): 91–5. doi:10.1016/0041-3879(80)90016-1. PMID 6776674.
- ↑ "WikiGenes - Collaborative Publishing". WikiGenes - Collaborative Publishing.
- ↑ Thompson MD, Capra V, Takasaki J, Maresca G, Rovati GE, Slutsky AS, Lilly C, Zamel N, McIntyre Burnham W, Cole DE, Siminovitch KA (2007). "A functional G300S variant of the cysteinyl leukotriene 1 receptor is associated with atopy in a Tristan da Cunha isolate". Pharmacogenetics and Genomics. 17 (7): 539–49. doi:10.1097/FPC.0b013e328012d0bf. PMID 17558309. S2CID 43781411.
- ↑ Yaddaden L, Véronneau S, Thompson MD, Rola-Pleszczynski M, Stankova J (2016). "Cellular signalling of cysteinyl leukotriene type 1 receptor variants CysLT₁-G300S and CysLT₁-I206S". Prostaglandins, Leukotrienes, and Essential Fatty Acids. 105: 1–8. doi:10.1016/j.plefa.2015.12.004. PMID 26869085.
- ↑ Kar M, Altıntoprak N, Muluk NB, Ulusoy S, Bafaqeeh SA, Cingi C (March 2016). "Antileukotrienes in adenotonsillar hypertrophy: a review of the literature". European Archives of Oto-Rhino-Laryngology. 273 (12): 4111–4117. doi:10.1007/s00405-016-3983-8. PMID 26980339. S2CID 31311115.
- ↑ Oussalah A, Mayorga C, Blanca M, Barbaud A, Nakonechna A, Cernadas J, Gotua M, Brockow K, Caubet JC, Bircher A, Atanaskovic M, Demoly P, K Tanno L, Terreehorst I, Laguna JJ, Romano A, Guéant JL (April 2016). "Genetic variants associated with drugs-induced immediate hypersensitivity reactions: a PRISMA-compliant systematic review". Allergy. 71 (4): 443–62. doi:10.1111/all.12821. PMID 26678823.
- ↑ Thompson MD, Takasaki J, Capra V, Rovati GE, Siminovitch KA, Burnham WM, Hudson TJ, Bossé Y, Cole DE (2006). "G-protein-coupled receptors and asthma endophenotypes: the cysteinyl leukotriene system in perspective". Molecular Diagnosis & Therapy. 10 (6): 353–66. doi:10.1007/bf03256212. PMID 17154652. S2CID 27541608.
- ↑ Szefler SJ, Phillips BR, Martinez FD, Chinchilli VM, Lemanske RF, Strunk RC, Zeiger RS, Larsen G, Spahn JD, Bacharier LB, Bloomberg GR, Guilbert TW, Heldt G, Morgan WJ, Moss MH, Sorkness CA, Taussig LM (2005). "Characterization of within-subject responses to fluticasone and montelukast in childhood asthma". The Journal of Allergy and Clinical Immunology. 115 (2): 233–42. doi:10.1016/j.jaci.2004.11.014. PMID 15696076.
Further reading
- Gronert K, Martinsson-Niskanen T, Ravasi S, Chiang N, Serhan CN (January 2001). "Selectivity of recombinant human leukotriene D(4), leukotriene B(4), and lipoxin A(4) receptors with aspirin-triggered 15-epi-LXA(4) and regulation of vascular and inflammatory responses". The American Journal of Pathology. 158 (1): 3–9. doi:10.1016/S0002-9440(10)63937-5. PMC 1850279. PMID 11141472.
- Sjöström M, Jakobsson PJ, Heimburger M, Palmblad J, Haeggström JZ (May 2001). "Human umbilical vein endothelial cells generate leukotriene C4 via microsomal glutathione S-transferase type 2 and express the CysLT(1) receptor". European Journal of Biochemistry. 268 (9): 2578–86. doi:10.1046/j.1432-1327.2001.02142.x. PMID 11322876.
- Mellor EA, Maekawa A, Austen KF, Boyce JA (July 2001). "Cysteinyl leukotriene receptor 1 is also a pyrimidinergic receptor and is expressed by human mast cells". Proceedings of the National Academy of Sciences of the United States of America. 98 (14): 7964–9. Bibcode:2001PNAS...98.7964M. doi:10.1073/pnas.141221498. PMC 35451. PMID 11438743.
- Mita H, Hasegawa M, Saito H, Akiyama K (November 2001). "Levels of cysteinyl leukotriene receptor mRNA in human peripheral leucocytes: significantly higher expression of cysteinyl leukotriene receptor 2 mRNA in eosinophils". Clinical and Experimental Allergy. 31 (11): 1714–23. doi:10.1046/j.1365-2222.2001.01184.x. PMID 11696047. S2CID 43278015.
- Shirasaki H, Kanaizumi E, Watanabe K, Matsui T, Sato J, Narita S, Rautiainen M, Himi T (July 2002). "Expression and localization of the cysteinyl leukotriene 1 receptor in human nasal mucosa". Clinical and Experimental Allergy. 32 (7): 1007–12. doi:10.1046/j.1365-2222.2002.01425.x. PMID 12100046. S2CID 25871662.
- Ohshima N, Nagase H, Koshino T, Miyamasu M, Yamaguchi M, Hirai K, Yamamoto K, Fujisawa T, Nakagawa N, Kishikawa K, Morita Y (September 2002). "A functional study on CysLT(1) receptors in human eosinophils". International Archives of Allergy and Immunology. 129 (1): 67–75. doi:10.1159/000065175. PMID 12373000. S2CID 11948102.
- Ohd JF, Nielsen CK, Campbell J, Landberg G, Löfberg H, Sjölander A (January 2003). "Expression of the leukotriene D4 receptor CysLT1, COX-2, and other cell survival factors in colorectal adenocarcinomas". Gastroenterology. 124 (1): 57–70. doi:10.1053/gast.2003.50011. PMID 12512030.
- Chibana K, Ishii Y, Asakura T, Fukuda T (April 2003). "Up-regulation of cysteinyl leukotriene 1 receptor by IL-13 enables human lung fibroblasts to respond to leukotriene C4 and produce eotaxin". Journal of Immunology. 170 (8): 4290–5. doi:10.4049/jimmunol.170.8.4290. PMID 12682264.
- Espinosa K, Bossé Y, Stankova J, Rola-Pleszczynski M (May 2003). "CysLT1 receptor upregulation by TGF-beta and IL-13 is associated with bronchial smooth muscle cell proliferation in response to LTD4". The Journal of Allergy and Clinical Immunology. 111 (5): 1032–40. doi:10.1067/mai.2003.1451. PMID 12743568.
- Walch L, Norel X, Gascard JP, Brink C (2003). "Arachidonic Acid Inhibits Cysteinyl-Leukotriene Receptor Activation in Human Pulmonary Vessels". Advances in Prostaglandin, Leukotriene, and other Bioactive Lipid Research. Advances in Experimental Medicine and Biology. Vol. 525. pp. 75–9. doi:10.1007/978-1-4419-9194-2_15. ISBN 978-1-4613-4831-3. PMID 12751740.
- Nielsen CK, Ohd JF, Wikström K, Massoumi R, Paruchuri S, Juhas M, Sjölander A (2003). "The Leukotriene Receptor CYSLT1 and 5- Lipoxygenase Are Upregulated in Colon Cancer". Advances in Prostaglandin, Leukotriene, and other Bioactive Lipid Research. Advances in Experimental Medicine and Biology. Vol. 525. pp. 201–4. doi:10.1007/978-1-4419-9194-2_43. ISBN 978-1-4613-4831-3. PMID 12751768.
- Mechiche H, Naline E, Candenas L, Pinto FM, Birembault P, Advenier C, Devillier P (July 2003). "Effects of cysteinyl leukotrienes in small human bronchus and antagonist activity of montelukast and its metabolites". Clinical and Experimental Allergy. 33 (7): 887–94. doi:10.1046/j.1365-2222.2003.01696.x. PMID 12859443. S2CID 37457415.
- Yang G, Haczku A, Chen H, Martin V, Galczenski H, Tomer Y, Van Besien CR, Evans JF, Panettieri RA, Funk CD, Van Beisen CR (May 2004). "Transgenic smooth muscle expression of the human CysLT1 receptor induces enhanced responsiveness of murine airways to leukotriene D4". American Journal of Physiology. Lung Cellular and Molecular Physiology. 286 (5): L992–1001. doi:10.1152/ajplung.00367.2003. PMID 15064240.
- Naik S, Billington CK, Pascual RM, Deshpande DA, Stefano FP, Kohout TA, Eckman DM, Benovic JL, Penn RB (March 2005). "Regulation of cysteinyl leukotriene type 1 receptor internalization and signaling". The Journal of Biological Chemistry. 280 (10): 8722–32. doi:10.1074/jbc.M413014200. PMID 15590629.
- Corrigan C, Mallett K, Ying S, Roberts D, Parikh A, Scadding G, Lee T (February 2005). "Expression of the cysteinyl leukotriene receptors cysLT(1) and cysLT(2) in aspirin-sensitive and aspirin-tolerant chronic rhinosinusitis". The Journal of Allergy and Clinical Immunology. 115 (2): 316–22. doi:10.1016/j.jaci.2004.10.051. PMID 15696087.
External links
- "Leukotriene Receptors: CysLT1". IUPHAR Database of Receptors and Ion Channels. International Union of Basic and Clinical Pharmacology. Archived from the original on 2016-03-03. Retrieved 2008-12-05.
- Overview of all the structural information available in the PDB for UniProt: Q9Y271 (Cysteinyl leukotriene receptor 1) at the PDBe-KB.
This article incorporates text from the United States National Library of Medicine, which is in the public domain.