| TCP-1/cpn60 chaperonin family | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
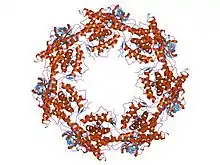 Structure of the bacterial chaperonin GroEL.[1] | |||||||||||
| Identifiers | |||||||||||
| Symbol | Cpn60_TCP1 | ||||||||||
| Pfam | PF00118 | ||||||||||
| InterPro | IPR002423 | ||||||||||
| PROSITE | PDOC00610 | ||||||||||
| CATH | 5GW5 | ||||||||||
| SCOP2 | 1grl / SCOPe / SUPFAM | ||||||||||
| CDD | cd00309 | ||||||||||
| |||||||||||
HSP60, also known as chaperonins (Cpn), is a family of heat shock proteins originally sorted by their 60kDa molecular mass. They prevent misfolding of proteins during stressful situations such as high heat, by assisting protein folding. HSP60 belong to a large class of molecules that assist protein folding, called molecular chaperones.[2][3]
Newly made proteins usually must fold from a linear chain of amino acids into a three-dimensional tertiary structure. The energy to fold proteins is supplied by non-covalent interactions between the amino acid side chains of each protein, and by solvent effects. Most proteins spontaneously fold into their most stable three-dimensional conformation, which is usually also their functional conformation, but occasionally proteins mis-fold. Molecular chaperones catalyze protein refolding by accelerating partial unfolding of misfolded proteins, aided by energy supplied by the hydrolysis of adenosine triphosphate (ATP). Chaperonin proteins may also tag misfolded proteins to be degraded.[3]
Structure
The structure of these chaperonins resemble two donuts stacked on top of one another to create a barrel. Each ring is composed of either 7, 8 or 9 subunits depending on the organism in which the chaperonin is found. Each ~60kDa peptide chain can be divided into three domains, apical, intermediate, and equatorial.[4]
The original chaperonin is proposed to have evolved from a peroxiredoxin.[5]
Classification
Group I
Group I chaperonins (Cpn60)[lower-alpha 1] are found in bacteria as well as organelles of endosymbiotic origin: chloroplasts and mitochondria.
The GroEL/GroES complex in E. coli is a Group I chaperonin and the best characterized large (~ 1 MDa) chaperonin complex.
- GroEL is a double-ring 14mer with a greasy hydrophobic patch at its opening and can accommodate the native folding of substrates 15-60 kDa in size.
- GroES (is a single-ring heptamer that binds to GroEL in the presence of ATP or transition state analogues of ATP hydrolysis, such as ADP-AlF3. It is like a cover that covers GroEL (box/bottle).
GroEL/GroES may not be able to undo protein aggregates, but kinetically it competes in the pathway of misfolding and aggregation, thereby preventing aggregate formation.[6]
The Cpn60 subfamily was discovered in 1988.[7] It was sequenced in 1992. The cpn10 and cpn60 oligomers also require Mg2+-ATP in order to interact to form a functional complex.[8] The binding of cpn10 to cpn60 inhibits the weak ATPase activity of cpn60.[9]
The RuBisCO subunit binding protein is a member of this family.[10] The crystal structure of Escherichia coli GroEL has been resolved to 2.8 Å.[11]
Some bacteria use multiple copies of this chaperonin, probably for different peptides.[4]
Group II

Group II chaperonins (TCP-1), found in the eukaryotic cytosol and in archaea, are more poorly characterized.
- The complex in archaea is called the thermosome. A homo-16mer in some archaea, it is regarded as the prototypical type II chaperonin.[lower-alpha 2]
- TRiC, the eukaryotic chaperonin, is composed of two rings of eight different though related subunits, each thought to be represented once per eight-membered ring. TRiC was originally thought to fold only the cytoskeletal proteins actin and tubulin but is now known to fold dozens of substrates.
Methanococcus maripaludis chaperonin (Mm cpn) is composed of sixteen identical subunits (eight per ring). It has been shown to fold the mitochondrial protein rhodanese; however, no natural substrates have yet been identified.[13]
Group II chaperonins are not thought to utilize a GroES-type cofactor to fold their substrates. They instead contain a "built-in" lid that closes in an ATP-dependent manner to encapsulate its substrates, a process that is required for optimal protein folding activity. They also interact with a co-chaperone, prefoldin, that helps move the substrate in.[3]
Other families
Group III includes some bacterial Cpns that are related to Group II. They have a lid, but the lid opening is noncooperative in them. They are thought to be an ancient relative of Group II.[3][4]
A Group I chaperonin gp146 from phage EL does not use a lid, and its donut interface is more similar to Group II. It might represent another ancient type of chaperonin.[14]
Mechanism of action
Chaperonins undergo large conformational changes during a folding reaction as a function of the enzymatic hydrolysis of ATP as well as binding of substrate proteins and cochaperonins, such as GroES. These conformational changes allow the chaperonin to bind an unfolded or misfolded protein, encapsulate that protein within one of the cavities formed by the two rings, and release the protein back into solution. Upon release, the substrate protein will either be folded or will require further rounds of folding, in which case it can again be bound by a chaperonin.
The exact mechanism by which chaperonins facilitate folding of substrate proteins is unknown. According to recent analyses by different experimental techniques, GroEL-bound substrate proteins populate an ensemble of compact and locally expanded states that lack stable tertiary interactions.[15] A number of models of chaperonin action have been proposed, which generally focus on two (not mutually exclusive) roles of chaperonin interior: passive and active. Passive models treat the chaperonin cage as an inert form, exerting influence by reducing the conformational space accessible to a protein substrate or preventing intermolecular interactions e.g. by aggregation prevention.[16] The active chaperonin role is in turn involved with specific chaperonin–substrate interactions that may be coupled to conformational rearrangements of the chaperonin.[17][18][19]
Probably the most popular model of the chaperonin active role is the iterative annealing mechanism (IAM), which focuses on the effect of iterative, and hydrophobic in nature, binding of the protein substrate to the chaperonin. According to computational simulation studies, the IAM leads to more productive folding by unfolding the substrate from misfolded conformations[19] or by prevention from protein misfolding through changing the folding pathway.[17]
Conservation of structural and functional homology
As mentioned, all cells contain chaperonins.
- In bacteria, the archetype is the well-characterized chaperonin GroEL from E. coli.
- In archaea, the chaperonin is called the thermosome.
- In eukarya, the cytoplasmic chaperonin is called CCT (also called TRiC).
These protein complexes appear to be essential for life in E. coli, Saccharomyces cerevisiae and higher eukaryotes. While there are differences between eukaryotic, bacterial and archaeal chaperonins, the general structure and mechanism are conserved.[3]
Bacteriophage T4 morphogenesis
The gene product 31 (gp31) of bacteriophage T4 is a protein required for bacteriophage morphogenesis that acts catalytically rather than being incorporated into the bacteriophage structure.[20] The bacterium E. coli is the host for bacteriophage T4. The bacteriophage encoded gp31 protein appears to be homologous to the E. coli cochaperonin protein GroES and is able to substitute for it in the assembly of phage T4 virions during infection.[21] Like GroES, gp31 forms a stable complex with GroEL chaperonin that is absolutely necessary for the folding and assembly in vivo of the bacteriophage T4 major capsid protein gp23.[21]
The main reason for the phage to need its own GroES homolog is that the gp23 protein is too large to fit into a conventional GroES cage. gp31 has longer loops that create a taller container.[22]
Clinical significance
Human GroEL is the immunodominant antigen of patients with Legionnaire's disease,[10] and is thought to play a role in the protection of the Legionella bacteria from oxygen radicals within macrophages. This hypothesis is based on the finding that the cpn60 gene is upregulated in response to hydrogen peroxide, a source of oxygen radicals. Cpn60 has also been found to display strong antigenicity in many bacterial species[23] and has the potential for inducing immune protection against unrelated bacterial infections.
Examples
Human genes encoding proteins containing this domain include:
See also
Notes
- ↑ The GroEL family is referred to, by InterPro, as Cpn60. However, CDD uses Cpn60 to refer to the Group II proteins in archaea.
- ↑ Some archaeons have evolved to use, like eukaryotes, different subunits. Methanosarcina acetivorans is known to have five types of subunits.[3] The ancestor to eukarotic TriC is thought to have two.[5]
References
- ↑ Braig K, Otwinowski Z, Hegde R, Boisvert DC, Joachimiak A, Horwich AL, Sigler PB (October 1994). "The crystal structure of the bacterial chaperonin GroEL at 2.8 A". Nature. 371 (6498): 578–86. Bibcode:1994Natur.371..578B. doi:10.1038/371578a0. PMID 7935790. S2CID 4341993.
- ↑ "Howard Hughes Investigators: Arthur L. Horwich, M.D." Archived from the original on 2019-07-26. Retrieved 2011-09-12.
- 1 2 3 4 5 6 Conway de Macario E, Yohda M, Macario AJ, Robb FT (2019-03-15). "Bridging human chaperonopathies and microbial chaperonins". Communications Biology. 2 (1): 103. doi:10.1038/s42003-019-0318-5. PMC 6420498. PMID 30911678.
- 1 2 3 Ansari MY, Mande SC (2018). "A Glimpse Into the Structure and Function of Atypical Type I Chaperonins". Frontiers in Molecular Biosciences. 5: 31. doi:10.3389/fmolb.2018.00031. PMC 5904260. PMID 29696145.
- 1 2 Willison, KR (5 October 2018). "The structure and evolution of eukaryotic chaperonin-containing TCP-1 and its mechanism that folds actin into a protein spring". The Biochemical Journal. 475 (19): 3009–3034. doi:10.1042/BCJ20170378. hdl:10044/1/63924. PMID 30291170. S2CID 52923821.
- ↑ Fenton WA, Horwich AL (May 2003). "Chaperonin-mediated protein folding: fate of substrate polypeptide". Quarterly Reviews of Biophysics. 36 (2): 229–56. doi:10.1017/S0033583503003883. PMID 14686103. S2CID 10328521.
- ↑ Hemmingsen SM, Woolford C, van der Vies SM, Tilly K, Dennis DT, Georgopoulos CP, et al. (May 1988). "Homologous plant and bacterial proteins chaperone oligomeric protein assembly". Nature. 333 (6171): 330–4. Bibcode:1988Natur.333..330H. doi:10.1038/333330a0. PMID 2897629. S2CID 4325057.
- ↑ Prasad TK, Stewart CR (March 1992). "cDNA clones encoding Arabidopsis thaliana and Zea mays mitochondrial chaperonin HSP60 and gene expression during seed germination and heat shock". Plant Molecular Biology. 18 (5): 873–85. doi:10.1007/BF00019202. PMID 1349837. S2CID 40768099.
- ↑ Schmidt A, Schiesswohl M, Völker U, Hecker M, Schumann W (June 1992). "Cloning, sequencing, mapping, and transcriptional analysis of the groESL operon from Bacillus subtilis". Journal of Bacteriology. 174 (12): 3993–9. doi:10.1128/jb.174.12.3993-3999.1992. PMC 206108. PMID 1350777.
- 1 2 Hindersson P, Høiby N, Bangsborg J (January 1991). "Sequence analysis of the Legionella micdadei groELS operon". FEMS Microbiology Letters. 61 (1): 31–8. doi:10.1111/j.1574-6968.1991.tb04317.x. PMID 1672279.
- ↑ Braig K, Otwinowski Z, Hegde R, Boisvert DC, Joachimiak A, Horwich AL, Sigler PB (October 1994). "The crystal structure of the bacterial chaperonin GroEL at 2.8 A". Nature. 371 (6498): 578–86. Bibcode:1994Natur.371..578B. doi:10.1038/371578a0. PMID 7935790. S2CID 4341993.
- ↑ Zang Y, Jin M, Wang H, Cui Z, Kong L, Liu C, Cong Y (December 2016). "Staggered ATP binding mechanism of eukaryotic chaperonin TRiC (CCT) revealed through high-resolution cryo-EM". Nature Structural & Molecular Biology. Springer Science and Business Media LLC. 23 (12): 1083–1091. doi:10.1038/nsmb.3309. PMID 27775711. S2CID 12001964.
- ↑ Kusmierczyk AR, Martin J (May 2003). "Nucleotide-dependent protein folding in the type II chaperonin from the mesophilic archaeon Methanococcus maripaludis". The Biochemical Journal. 371 (Pt 3): 669–73. doi:10.1042/BJ20030230. PMC 1223359. PMID 12628000.
- ↑ Bracher A, Paul SS, Wang H, Wischnewski N, Hartl FU, Hayer-Hartl M (27 April 2020). "Structure and conformational cycle of a bacteriophage-encoded chaperonin". PLOS ONE. 15 (4): e0230090. Bibcode:2020PLoSO..1530090B. doi:10.1371/journal.pone.0230090. PMC 7185714. PMID 32339190.
- ↑ Hartl FU, Hayer-Hartl M (June 2009). "Converging concepts of protein folding in vitro and in vivo". Nature Structural & Molecular Biology. 16 (6): 574–81. doi:10.1038/nsmb.1591. PMID 19491934. S2CID 205522841.
- ↑ Apetri AC, Horwich AL (November 2008). "Chaperonin chamber accelerates protein folding through passive action of preventing aggregation". Proceedings of the National Academy of Sciences of the United States of America. 105 (45): 17351–5. Bibcode:2008PNAS..10517351A. doi:10.1073/pnas.0809794105. PMC 2579888. PMID 18987317.
- 1 2 Kmiecik S, Kolinski A (July 2011). "Simulation of chaperonin effect on protein folding: a shift from nucleation-condensation to framework mechanism". Journal of the American Chemical Society. 133 (26): 10283–9. doi:10.1021/ja203275f. PMC 3132998. PMID 21618995.
- ↑ Chakraborty K, Chatila M, Sinha J, Shi Q, Poschner BC, Sikor M, et al. (July 2010). "Chaperonin-catalyzed rescue of kinetically trapped states in protein folding". Cell. 142 (1): 112–22. doi:10.1016/j.cell.2010.05.027. PMID 20603018. S2CID 3859016.
- 1 2 Todd MJ, Lorimer GH, Thirumalai D (April 1996). "Chaperonin-facilitated protein folding: optimization of rate and yield by an iterative annealing mechanism". Proceedings of the National Academy of Sciences of the United States of America. 93 (9): 4030–5. Bibcode:1996PNAS...93.4030T. doi:10.1073/pnas.93.9.4030. PMC 39481. PMID 8633011.
- ↑ Snustad DP (August 1968). "Dominance interactions in Escherichia coli cells mixedly infected with bacteriophage T4D wild-type and amber mutants and their possible implications as to type of gene-product function: catalytic vs. stoichiometric". Virology. 35 (4): 550–63. doi:10.1016/0042-6822(68)90285-7. PMID 4878023.
- 1 2 Marusich EI, Kurochkina LP, Mesyanzhinov VV (April 1998). "Chaperones in bacteriophage T4 assembly". Biochemistry (Moscow). 63 (4): 399–406. PMID 9556522.
- ↑ Bukau B, Horwich AL (February 1998). "The Hsp70 and Hsp60 chaperone machines". Cell. 92 (3): 351–66. doi:10.1016/S0092-8674(00)80928-9. PMID 9476895. S2CID 16526409.
- ↑ Gor D, Mayfield JE (February 1992). "Cloning and nucleotide sequence of the Brucella abortus groE operon". Biochimica et Biophysica Acta (BBA) - Gene Structure and Expression. 1130 (1): 120–2. doi:10.1016/0167-4781(92)90476-g. PMID 1347461.
External links
- more details... Archived 2011-03-01 at the Wayback Machine
- Chaperonins at the U.S. National Library of Medicine Medical Subject Headings (MeSH)
- cpnDB: a chaperonin database
- Animations of activity of Chaperonins
- NIH Material on HSP60
- HSP60 Archived 2008-03-16 at the Wayback Machine
- HSP60 in Flies
- The Chaperonin Home Page
- The Protein Data Bank
- Enzyme Database on HSP60 Archived 2021-04-18 at the Wayback Machine
- HSP60 on Pub Med
- HSP60 Gene Report
- HSP60+Heat-Shock+Proteins at the U.S. National Library of Medicine Medical Subject Headings (MeSH)