.png.webp)


Forkhead box protein P2 (FOXP2) is a protein that, in humans, is encoded by the FOXP2 gene. FOXP2 is a member of the forkhead box family of transcription factors, proteins that regulate gene expression by binding to DNA. It is expressed in the brain, heart, lungs and digestive system.[3][4]
FOXP2 is found in many vertebrates, where it plays an important role in mimicry in birds (such as birdsong) and echolocation in bats. FOXP2 is also required for the proper development of speech and language in humans.[5] In humans, mutations in FOXP2 cause the severe speech and language disorder developmental verbal dyspraxia.[5][6] Studies of the gene in mice and songbirds indicate that it is necessary for vocal imitation and the related motor learning.[7][8][9] Outside the brain, FOXP2 has also been implicated in development of other tissues such as the lung and digestive system.[10]
Initially identified in 1998 as the genetic cause of a speech disorder in a British family designated the KE family, FOXP2 was the first gene discovered to be associated with speech and language[11] and was subsequently dubbed "the language gene".[12] However, other genes are necessary for human language development, and a 2018 analysis confirmed that there was no evidence of recent positive evolutionary selection of FOXP2 in humans.[13][14]
Structure and function

As a FOX protein, FOXP2 contains a forkhead-box domain. In addition, it contains a polyglutamine tract, a zinc finger and a leucine zipper. The protein attaches to the DNA of other proteins and controls their activity through the forkhead-box domain. Only a few targeted genes have been identified, however researchers believe that there could be up to hundreds of other genes targeted by the FOXP2 gene. The forkhead box P2 protein is active in the brain and other tissues before and after birth, many studies show that it is paramount for the growth of nerve cells and transmission between them. The FOXP2 gene is also involved in synaptic plasticity, making it imperative for learning and memory.[15]
FOXP2 is required for proper brain and lung development. Knockout mice with only one functional copy of the FOXP2 gene have significantly reduced vocalizations as pups.[16] Knockout mice with no functional copies of FOXP2 are runted, display abnormalities in brain regions such as the Purkinje layer, and die an average of 21 days after birth from inadequate lung development.[10]
FOXP2 is expressed in many areas of the brain,[17] including the basal ganglia and inferior frontal cortex, where it is essential for brain maturation and speech and language development.[18] In mice, the gene was found to be twice as highly expressed in male pups than female pups, which correlated with an almost double increase in the number of vocalisations the male pups made when separated from mothers. Conversely, in human children aged 4–5, the gene was found to be 30% more expressed in the Broca's areas of female children. The researchers suggested that the gene is more active in "the more communicative sex".[19][20]
The expression of FOXP2 is subject to post-transcriptional regulation, particularly microRNA (miRNA), causing the repression of the FOXP2 3' untranslated region.[21]
Three amino acid substitutions distinguish the human FOXP2 protein from that found in mice, while two amino acid substitutions distinguish the human FOXP2 protein from that found in chimpanzees,[17] but only one of these changes is unique to humans.[10] Evidence from genetically manipulated mice[22] and human neuronal cell models[23] suggests that these changes affect the neural functions of FOXP2.
Clinical significance
The FOXP2 gene has been implicated in several cognitive functions including; general brain development, language, and synaptic plasticity. The FOXP2 gene region acts as a transcription factor for the forkhead box P2 protein. Transcription factors affect other regions, and the forkhead box P2 protein has been suggested to also act as a transcription factor for hundreds of genes. This prolific involvement opens the possibility that the FOXP2 gene is much more extensive than originally thought.[15] Other targets of transcription have been researched without correlation to FOXP2. Specifically, FOXP2 has been investigated in correlation with autism and dyslexia, however with no mutation was discovered as the cause.[24][6] One well identified target is language.[25] Although some research disagrees with this correlation,[26] the majority of research shows that a mutated FOXP2 causes the observed production deficiency.[15][25][27][24][28][29]
There is some evidence that the linguistic impairments associated with a mutation of the FOXP2 gene are not simply the result of a fundamental deficit in motor control. Brain imaging of affected individuals indicates functional abnormalities in language-related cortical and basal ganglia regions, demonstrating that the problems extend beyond the motor system.[30]
Mutations in FOXP2 are among several (26 genes plus 2 intergenic) loci which correlate to ADHD diagnosis in adults – clinical ADHD is an umbrella label for a heterogeneous group of genetic and neurological phenomena which may result from FOXP2 mutations or other causes.[31]
A 2020 genome-wide association study (GWAS) implicates single-nucleotide polymorphisms (SNPs) of FOXP2 in susceptibility to cannabis use disorder.[32]
Language disorder
It is theorized that the translocation of the 7q31.2 region of the FOXP2 gene causes a severe language impairment called developmental verbal dyspraxia (DVD)[25] or childhood apraxia of speech (CAS)[33] So far this type of mutation has only been discovered in three families across the world including the original KE family.[29] A missense mutation causing an arginine-to-histidine substitution (R553H) in the DNA-binding domain is thought to be the abnormality in KE.[34] This would cause a normally basic residue to be fairly acidic and highly reactive at the body's pH. A heterozygous nonsense mutation, R328X variant, produces a truncated protein involved in speech and language difficulties in one KE individual and two of their close family members. R553H and R328X mutations also affected nuclear localization, DNA-binding, and the transactivation (increased gene expression) properties of FOXP2.[6]
These individuals present with deletions, translocations, and missense mutations. When tasked with repetition and verb generation, these individuals with DVD/CAS had decreased activation in the putamen and Broca's area in fMRI studies. These areas are commonly known as areas of language function.[35] This is one of the primary reasons that FOXP2 is known as a language gene. They have delayed onset of speech, difficulty with articulation including slurred speech, stuttering, and poor pronunciation, as well as dyspraxia.[29] It is believed that a major part of this speech deficit comes from an inability to coordinate the movements necessary to produce normal speech including mouth and tongue shaping.[25] Additionally, there are more general impairments with the processing of the grammatical and linguistic aspects of speech.[6] These findings suggest that the effects of FOXP2 are not limited to motor control, as they include comprehension among other cognitive language functions. General mild motor and cognitive deficits are noted across the board.[27] Clinically these patients can also have difficulty coughing, sneezing, or clearing their throats.[25]
While FOXP2 has been proposed to play a critical role in the development of speech and language, this view has been challenged by the fact that the gene is also expressed in other mammals as well as birds and fish that do not speak.[36] It has also been proposed that the FOXP2 transcription-factor is not so much a hypothetical 'language gene' but rather part of a regulatory machinery related to externalization of speech.[37]
Evolution

The FOXP2 gene is highly conserved in mammals.[17] The human gene differs from that in non-human primates by the substitution of two amino acids, a threonine to asparagine substitution at position 303 (T303N) and an asparagine to serine substitution at position 325 (N325S).[34] In mice it differs from that of humans by three substitutions, and in zebra finch by seven amino acids.[17][38][39] One of the two amino acid differences between human and chimps also arose independently in carnivores and bats.[10][40] Similar FOXP2 proteins can be found in songbirds, fish, and reptiles such as alligators.[41][42]
DNA sampling from Homo neanderthalensis bones indicates that their FOXP2 gene is a little different though largely similar to those of Homo sapiens (i.e. humans).[43][44] Previous genetic analysis had suggested that the H. sapiens FOXP2 gene became fixed in the population around 125,000 years ago.[45] Some researchers consider the Neanderthal findings to indicate that the gene instead swept through the population over 260,000 years ago, before our most recent common ancestor with the Neanderthals.[45] Other researchers offer alternative explanations for how the H. sapiens version would have appeared in Neanderthals living 43,000 years ago.[45]
According to a 2002 study, the FOXP2 gene showed indications of recent positive selection.[17][46] Some researchers have speculated that positive selection is crucial for the evolution of language in humans.[17] Others, however, were unable to find a clear association between species with learned vocalizations and similar mutations in FOXP2.[41][42] A 2018 analysis of a large sample of globally distributed genomes confirmed there was no evidence of positive selection, suggesting that the original signal of positive selection may be driven by sample composition.[13][14] Insertion of both human mutations into mice, whose version of FOXP2 otherwise differs from the human and chimpanzee versions in only one additional base pair, causes changes in vocalizations as well as other behavioral changes, such as a reduction in exploratory tendencies, and a decrease in maze learning time. A reduction in dopamine levels and changes in the morphology of certain nerve cells are also observed.[22]
Interactions
FOXP2 is known to regulate CNTNAP2, CTBP1,[47] SRPX2 and SCN3A.[48][18][49]
FOXP2 downregulates CNTNAP2, a member of the neurexin family found in neurons. CNTNAP2 is associated with common forms of language impairment.[50]
FOXP2 also downregulates SRPX2, the 'Sushi Repeat-containing Protein X-linked 2'.[51][52] It directly reduces its expression, by binding to its gene's promoter. SRPX2 is involved in glutamatergic synapse formation in the cerebral cortex and is more highly expressed in childhood. SRPX2 appears to specifically increase the number of glutamatergic synapses in the brain, while leaving inhibitory GABAergic synapses unchanged and not affecting dendritic spine length or shape. On the other hand, FOXP2's activity does reduce dendritic spine length and shape, in addition to number, indicating it has other regulatory roles in dendritic morphology.[51]
In other animals
Chimpanzees
In chimpanzees, FOXP2 differs from the human version by two amino acids.[53] A study in Germany sequenced FOXP2's complementary DNA in chimps and other species to compare it with human complementary DNA in order to find the specific changes in the sequence.[17] FOXP2 was found to be functionally different in humans compared to chimps. Since FOXP2 was also found to have an effect on other genes, its effects on other genes is also being studied.[54] Researchers deduced that there could also be further clinical applications in the direction of these studies in regards to illnesses that show effects on human language ability.[23]
Mice
In a mouse FOXP2 gene knockouts, loss of both copies of the gene causes severe motor impairment related to cerebellar abnormalities and lack of ultrasonic vocalisations normally elicited when pups are removed from their mothers.[16] These vocalizations have important communicative roles in mother–offspring interactions. Loss of one copy was associated with impairment of ultrasonic vocalisations and a modest developmental delay. Male mice on encountering female mice produce complex ultrasonic vocalisations that have characteristics of song.[55] Mice that have the R552H point mutation carried by the KE family show cerebellar reduction and abnormal synaptic plasticity in striatal and cerebellar circuits.[7]
Humanized FOXP2 mice display altered cortico-basal ganglia circuits. The human allele of the FOXP2 gene was transferred into the mouse embryos through homologous recombination to create humanized FOXP2 mice. The human variant of FOXP2 also had an effect on the exploratory behavior of the mice. In comparison to knockout mice with one non-functional copy of FOXP2, the humanized mouse model showed opposite effects when testing its effect on the levels of dopamine, plasticity of synapses, patterns of expression in the striatum and behavior that was exploratory in nature.[22]
When FOXP2 expression was altered in mice, it affected many different processes including the learning motor skills and the plasticity of synapses. Additionally, FOXP2 is found more in the sixth layer of the cortex than in the fifth, and this is consistent with it having greater roles in sensory integration. FOXP2 was also found in the medial geniculate nucleus of the mouse brain, which is the processing area that auditory inputs must go through in the thalamus. It was found that its mutations play a role in delaying the development of language learning. It was also found to be highly expressed in the Purkinje cells and cerebellar nuclei of the cortico-cerebellar circuits. High FOXP2 expression has also been shown in the spiny neurons that express type 1 dopamine receptors in the striatum, substantia nigra, subthalamic nucleus and ventral tegmental area. The negative effects of the mutations of FOXP2 in these brain regions on motor abilities were shown in mice through tasks in lab studies. When analyzing the brain circuitry in these cases, scientists found greater levels of dopamine and decreased lengths of dendrites, which caused defects in long-term depression, which is implicated in motor function learning and maintenance. Through EEG studies, it was also found that these mice had increased levels of activity in their striatum, which contributed to these results. There is further evidence for mutations of targets of the FOXP2 gene shown to have roles in schizophrenia, epilepsy, autism, bipolar disorder and intellectual disabilities.[56]
Bats
FOXP2 has implications in the development of bat echolocation.[34][40][57] Contrary to apes and mice, FOXP2 is extremely diverse in echolocating bats.[40] Twenty-two sequences of non-bat eutherian mammals revealed a total number of 20 nonsynonymous mutations in contrast to half that number of bat sequences, which showed 44 nonsynonymous mutations.[40] All cetaceans share three amino acid substitutions, but no differences were found between echolocating toothed whales and non-echolocating baleen cetaceans.[40] Within bats, however, amino acid variation correlated with different echolocating types.[40]
Birds
In songbirds, FOXP2 most likely regulates genes involved in neuroplasticity.[8][58] Gene knockdown of FOXP2 in area X of the basal ganglia in songbirds results in incomplete and inaccurate song imitation.[8] Overexpression of FOXP2 was accomplished through injection of adeno-associated virus serotype 1 (AAV1) into area X of the brain. This overexpression produced similar effects to that of knockdown; juvenile zebra finch birds were unable to accurately imitate their tutors.[59] Similarly, in adult canaries, higher FOXP2 levels also correlate with song changes.[39]
Levels of FOXP2 in adult zebra finches are significantly higher when males direct their song to females than when they sing song in other contexts.[58] "Directed" singing refers to when a male is singing to a female usually for a courtship display. "Undirected" singing occurs when for example, a male sings when other males are present or is alone.[60] Studies have found that FoxP2 levels vary depending on the social context. When the birds were singing undirected song, there was a decrease of FoxP2 expression in Area X. This downregulation was not observed and FoxP2 levels remained stable in birds singing directed song.[58]
Differences between song-learning and non-song-learning birds have been shown to be caused by differences in FOXP2 gene expression, rather than differences in the amino acid sequence of the FOXP2 protein.
Zebrafish
In zebrafish, FOXP2 is expressed in the ventral and dorsal thalamus, telencephalon, diencephalon where it likely plays a role in nervous system development. The zebrafish FOXP2 gene has an 85% similarity to the human FOX2P ortholog.[61]
History
FOXP2 and its gene were discovered as a result of investigations on an English family known as the KE family, half of whom (15 individuals across three generations) had a speech and language disorder called developmental verbal dyspraxia. Their case was studied at the Institute of Child Health of University College London.[62] In 1990, Myrna Gopnik, Professor of Linguistics at McGill University, reported that the disorder-affected KE family had severe speech impediment with incomprehensible talk, largely characterized by grammatical deficits.[63] She hypothesized that the basis was not of learning or cognitive disability, but due to genetic factors affecting mainly grammatical ability.[64] (Her hypothesis led to a popularised existence of "grammar gene" and a controversial notion of grammar-specific disorder.[65][66]) In 1995, the University of Oxford and the Institute of Child Health researchers found that the disorder was purely genetic.[67] Remarkably, the inheritance of the disorder from one generation to the next was consistent with autosomal dominant inheritance, i.e., mutation of only a single gene on an autosome (non-sex chromosome) acting in a dominant fashion. This is one of the few known examples of Mendelian (monogenic) inheritance for a disorder affecting speech and language skills, which typically have a complex basis involving multiple genetic risk factors.[68]

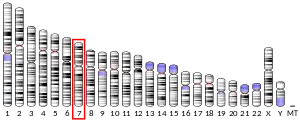
In 1998, Oxford University geneticists Simon Fisher, Anthony Monaco, Cecilia S. L. Lai, Jane A. Hurst, and Faraneh Vargha-Khadem identified an autosomal dominant monogenic inheritance that is localized on a small region of chromosome 7 from DNA samples taken from the affected and unaffected members.[3] The chromosomal region (locus) contained 70 genes.[69] The locus was given the official name "SPCH1" (for speech-and-language-disorder-1) by the Human Genome Nomenclature committee. Mapping and sequencing of the chromosomal region was performed with the aid of bacterial artificial chromosome clones.[4] Around this time, the researchers identified an individual who was unrelated to the KE family but had a similar type of speech and language disorder. In this case, the child, known as CS, carried a chromosomal rearrangement (a translocation) in which part of chromosome 7 had become exchanged with part of chromosome 5. The site of breakage of chromosome 7 was located within the SPCH1 region.[4]
In 2001, the team identified in CS that the mutation is in the middle of a protein-coding gene.[5] Using a combination of bioinformatics and RNA analyses, they discovered that the gene codes for a novel protein belonging to the forkhead-box (FOX) group of transcription factors. As such, it was assigned with the official name of FOXP2. When the researchers sequenced the FOXP2 gene in the KE family, they found a heterozygous point mutation shared by all the affected individuals, but not in unaffected members of the family and other people.[5] This mutation is due to an amino-acid substitution that inhibits the DNA-binding domain of the FOXP2 protein.[70] Further screening of the gene identified multiple additional cases of FOXP2 disruption, including different point mutations[6] and chromosomal rearrangements,[71] providing evidence that damage to one copy of this gene is sufficient to derail speech and language development.
See also
References
- 1 2 3 GRCh38: Ensembl release 89: ENSG00000128573 - Ensembl, May 2017
- ↑ "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- 1 2 Fisher SE, Vargha-Khadem F, Watkins KE, Monaco AP, Pembrey ME (February 1998). "Localisation of a gene implicated in a severe speech and language disorder". Nature Genetics. 18 (2): 168–70. doi:10.1038/ng0298-168. hdl:11858/00-001M-0000-0012-CBD9-5. PMID 9462748. S2CID 3190318.
- 1 2 3 Lai CS, Fisher SE, Hurst JA, Levy ER, Hodgson S, Fox M, et al. (August 2000). "The SPCH1 region on human 7q31: genomic characterization of the critical interval and localization of translocations associated with speech and language disorder". American Journal of Human Genetics. 67 (2): 357–68. doi:10.1086/303011. PMC 1287211. PMID 10880297.
- 1 2 3 4 Lai CS, Fisher SE, Hurst JA, Vargha-Khadem F, Monaco AP (October 2001). "A forkhead-domain gene is mutated in a severe speech and language disorder". Nature. 413 (6855): 519–23. Bibcode:2001Natur.413..519L. doi:10.1038/35097076. hdl:11858/00-001M-0000-0012-CB9C-F. PMID 11586359. S2CID 4421562.
- 1 2 3 4 5 MacDermot KD, Bonora E, Sykes N, Coupe AM, Lai CS, Vernes SC, et al. (June 2005). "Identification of FOXP2 truncation as a novel cause of developmental speech and language deficits". American Journal of Human Genetics. 76 (6): 1074–80. doi:10.1086/430841. PMC 1196445. PMID 15877281.
- 1 2 Groszer M, Keays DA, Deacon RM, de Bono JP, Prasad-Mulcare S, Gaub S, et al. (March 2008). "Impaired synaptic plasticity and motor learning in mice with a point mutation implicated in human speech deficits". Current Biology. 18 (5): 354–62. doi:10.1016/j.cub.2008.01.060. PMC 2917768. PMID 18328704.
- 1 2 3 Haesler S, Rochefort C, Georgi B, Licznerski P, Osten P, Scharff C (December 2007). "Incomplete and inaccurate vocal imitation after knockdown of FoxP2 in songbird basal ganglia nucleus Area X". PLOS Biology. 5 (12): e321. doi:10.1371/journal.pbio.0050321. PMC 2100148. PMID 18052609.
- ↑ Fisher SE, Scharff C (April 2009). "FOXP2 as a molecular window into speech and language". Trends in Genetics. 25 (4): 166–77. doi:10.1016/j.tig.2009.03.002. hdl:11858/00-001M-0000-0012-CA31-7. PMID 19304338.
- 1 2 3 4 Shu W, Lu MM, Zhang Y, Tucker PW, Zhou D, Morrisey EE (May 2007). "Foxp2 and Foxp1 cooperatively regulate lung and esophagus development". Development. 134 (10): 1991–2000. doi:10.1242/dev.02846. PMID 17428829. S2CID 22896384.
- ↑ Nudel R, Newbury DF (September 2013). "FOXP2". Wiley Interdisciplinary Reviews. Cognitive Science. 4 (5): 547–560. doi:10.1002/wcs.1247. PMC 3992897. PMID 24765219.
- ↑ Harpaz Y. "Language gene found". human-brain.org. Archived from the original on 25 October 2014. Retrieved 31 October 2014.
- 1 2 Atkinson EG, Audesse AJ, Palacios JA, Bobo DM, Webb AE, Ramachandran S, Henn BM (September 2018). "No Evidence for Recent Selection at FOXP2 among Diverse Human Populations". Cell. 174 (6): 1424–1435.e15. doi:10.1016/j.cell.2018.06.048. PMC 6128738. PMID 30078708.
- 1 2 "Language Gene Dethroned". The Scientist Magazine®. Retrieved 28 January 2020.
- 1 2 3 "FOXP2 Gene". Genetics Home Reference. U.S. National Library of Medicine, National Institutes of Health. September 2016.
- 1 2 Shu W, Cho JY, Jiang Y, Zhang M, Weisz D, Elder GA, et al. (July 2005). "Altered ultrasonic vocalization in mice with a disruption in the Foxp2 gene". Proceedings of the National Academy of Sciences of the United States of America. 102 (27): 9643–8. Bibcode:2005PNAS..102.9643S. doi:10.1073/pnas.0503739102. PMC 1160518. PMID 15983371.
- 1 2 3 4 5 6 7 Enard W, Przeworski M, Fisher SE, Lai CS, Wiebe V, Kitano T, et al. (August 2002). "Molecular evolution of FOXP2, a gene involved in speech and language" (PDF). Nature. 418 (6900): 869–72. Bibcode:2002Natur.418..869E. doi:10.1038/nature01025. hdl:11858/00-001M-0000-0012-CB89-A. PMID 12192408. S2CID 4416233. Archived from the original (PDF) on 30 August 2006.
- 1 2 Spiteri E, Konopka G, Coppola G, Bomar J, Oldham M, Ou J, et al. (December 2007). "Identification of the transcriptional targets of FOXP2, a gene linked to speech and language, in developing human brain". American Journal of Human Genetics. 81 (6): 1144–57. doi:10.1086/522237. PMC 2276350. PMID 17999357.
- ↑ Balter M (February 2013). "'Language Gene' More Active in Young Girls Than Boy". Science: 360.
- ↑ Bowers JM, Perez-Pouchoulen M, Edwards NS, McCarthy MM (February 2013). "Foxp2 mediates sex differences in ultrasonic vocalization by rat pups and directs order of maternal retrieval". The Journal of Neuroscience. 33 (8): 3276–83. doi:10.1523/JNEUROSCI.0425-12.2013. PMC 3727442. PMID 23426656.
- ↑ Clovis YM, Enard W, Marinaro F, Huttner WB, De Pietri Tonelli D (September 2012). "Convergent repression of Foxp2 3'UTR by miR-9 and miR-132 in embryonic mouse neocortex: implications for radial migration of neurons". Development. 139 (18): 3332–42. doi:10.1242/dev.078063. PMID 22874921.
- 1 2 3 Enard W, Gehre S, Hammerschmidt K, Hölter SM, Blass T, Somel M, et al. (May 2009). "A humanized version of Foxp2 affects cortico-basal ganglia circuits in mice". Cell. 137 (5): 961–71. doi:10.1016/j.cell.2009.03.041. hdl:11858/00-001M-0000-000F-F8C5-2. PMID 19490899. S2CID 667723.
- 1 2 Konopka G, Bomar JM, Winden K, Coppola G, Jonsson ZO, Gao F, et al. (November 2009). "Human-specific transcriptional regulation of CNS development genes by FOXP2". Nature. 462 (7270): 213–7. Bibcode:2009Natur.462..213K. doi:10.1038/nature08549. PMC 2778075. PMID 19907493.
- "Why Can't Chimps Speak? Key Differences In How Human And Chimp Versions Of FOXP2 Gene Work". ScienceDaily (Press release). 12 November 2009.
- 1 2 Gauthier J, Joober R, Mottron L, Laurent S, Fuchs M, De Kimpe V, Rouleau GA (April 2003). "Mutation screening of FOXP2 in individuals diagnosed with autistic disorder". American Journal of Medical Genetics. Part A. 118A (2): 172–5. doi:10.1002/ajmg.a.10105. PMID 12655497. S2CID 39762074.
- 1 2 3 4 5 "FOXP2-related speech and language disorder". Genetics Home Reference. U.S. National Library of Medicine, National Institutes of Health. Retrieved 26 February 2019.
- ↑ Newbury DF, Bonora E, Lamb JA, Fisher SE, Lai CS, Baird G, et al. (May 2002). "FOXP2 is not a major susceptibility gene for autism or specific language impairment". American Journal of Human Genetics. 70 (5): 1318–27. doi:10.1086/339931. PMC 447606. PMID 11894222.
- 1 2 Lennon PA, Cooper ML, Peiffer DA, Gunderson KL, Patel A, Peters S, et al. (April 2007). "Deletion of 7q31.1 supports involvement of FOXP2 in language impairment: clinical report and review". American Journal of Medical Genetics. Part A. 143A (8): 791–8. doi:10.1002/ajmg.a.31632. PMID 17330859. S2CID 22021740.
- ↑ Rossell S, Tan E, Bozaoglu K, Neill E, Sumner P, Carruthers S, Van Rheenen T, Thomas E, Gurvich C (2017). "Is language Impairment in Schizophrenia related to Language Genes?". European Neuropsychopharmacology. 27: S459–S460. doi:10.1016/j.euroneuro.2016.09.532. S2CID 54316143.
- 1 2 3 Reuter MS, Riess A, Moog U, Briggs TA, Chandler KE, Rauch A, et al. (January 2017). "FOXP2 variants in 14 individuals with developmental speech and language disorders broaden the mutational and clinical spectrum". Journal of Medical Genetics. 54 (1): 64–72. doi:10.1136/jmedgenet-2016-104094. PMID 27572252. S2CID 24589445.
- ↑ Liégeois F, Baldeweg T, Connelly A, Gadian DG, Mishkin M, Vargha-Khadem F (November 2003). "Language fMRI abnormalities associated with FOXP2 gene mutation". Nature Neuroscience. 6 (11): 1230–7. doi:10.1038/nn1138. PMID 14555953. S2CID 31003547.
- ↑ Demontis D, Walters RK, Martin J, Mattheisen M, Als TD, Agerbo E, et al. (January 2019). "Discovery of the first genome-wide significant risk loci for attention deficit/hyperactivity disorder". Nature Genetics. 51 (1): 63–75. doi:10.1038/s41588-018-0269-7. PMC 6481311. PMID 30478444.
- ↑ Johnson EC, Demontis D, Thorgeirsson TE, Walters RK, Polimanti R, Hatoum AS, et al. (December 2020). "A large-scale genome-wide association study meta-analysis of cannabis use disorder". The Lancet. Psychiatry. 7 (12): 1032–1045. doi:10.1016/S2215-0366(20)30339-4. PMC 7674631. PMID 33096046.
- ↑ Morgan A, Fisher SE, Scheffer I, Hildebrand M (23 June 2016). Adam MP, Ardinger HH, Pagon RA, Wallace SE, Bean LJ, Stephens K, Amemiya A (eds.). "FOXP2-Related Speech and Language Disorders". GenReviews. University of Washington. PMID 27336128.
- 1 2 3 Preuss TM (June 2012). "Human brain evolution: from gene discovery to phenotype discovery". Proceedings of the National Academy of Sciences of the United States of America. 109 (Suppl 1): 10709–16. Bibcode:2012PNAS..10910709P. doi:10.1073/pnas.1201894109. PMC 3386880. PMID 22723367.
- ↑ Vargha-Khadem F, Gadian DG, Copp A, Mishkin M (February 2005). "FOXP2 and the neuroanatomy of speech and language". Nature Reviews. Neuroscience. 6 (2): 131–8. doi:10.1038/nrn1605. PMID 15685218. S2CID 2504002.
- ↑ Friederici AD (2016). 'Language in the Brain. Cambridge, MA: The MIT Press. p. 222. ISBN 978-0-262-03692-4.
- ↑ Berwick RC, Chomsky N (2016). Why Only Us?. Cambridge, MA: The MIT Press. p. 76. ISBN 978-0-262-53349-2.
- ↑ Teramitsu I, Kudo LC, London SE, Geschwind DH, White SA (March 2004). "Parallel FoxP1 and FoxP2 expression in songbird and human brain predicts functional interaction". The Journal of Neuroscience. 24 (13): 3152–63. doi:10.1523/JNEUROSCI.5589-03.2004. PMC 6730014. PMID 15056695.
- 1 2 Haesler S, Wada K, Nshdejan A, Morrisey EE, Lints T, Jarvis ED, Scharff C (March 2004). "FoxP2 expression in avian vocal learners and non-learners". The Journal of Neuroscience. 24 (13): 3164–75. doi:10.1523/JNEUROSCI.4369-03.2004. PMC 6730012. PMID 15056696.
- 1 2 3 4 5 6 Li G, Wang J, Rossiter SJ, Jones G, Zhang S (September 2007). Ellegren H (ed.). "Accelerated FoxP2 evolution in echolocating bats". PLOS ONE. 2 (9): e900. Bibcode:2007PLoSO...2..900L. doi:10.1371/journal.pone.0000900. PMC 1976393. PMID 17878935.
- 1 2 Webb DM, Zhang J (2005). "FoxP2 in song-learning birds and vocal-learning mammals". The Journal of Heredity. 96 (3): 212–6. doi:10.1093/jhered/esi025. PMID 15618302.
- 1 2 Scharff C, Haesler S (December 2005). "An evolutionary perspective on FoxP2: strictly for the birds?". Current Opinion in Neurobiology. 15 (6): 694–703. doi:10.1016/j.conb.2005.10.004. PMID 16266802. S2CID 11350165.
- ↑ Zimmer C (17 March 2016). "Humans Interbred With Hominins on Multiple Occasions, Study Finds". The New York Times. Retrieved 17 March 2016.
- ↑ Krause J, Lalueza-Fox C, Orlando L, Enard W, Green RE, Burbano HA, et al. (November 2007). "The derived FOXP2 variant of modern humans was shared with Neandertals". Current Biology. 17 (21): 1908–12. doi:10.1016/j.cub.2007.10.008. hdl:11858/00-001M-0000-000F-FED3-1. PMID 17949978. S2CID 9518208.
- Nicholas Wade (19 October 2007). "Neanderthals Had Important Speech Gene, DNA Evidence Shows". The New York Times.
- 1 2 3 Benítez-Burraco A, Longa VM, Lorenzo G, Uriagereka J (November 2008). "Also sprach Neanderthalis... Or Did She?". Biolinguistics. 2 (2–3): 225–232. doi:10.5964/bioling.8643. S2CID 60864520.
- ↑ Toda M, Okubo S, Ikigai H, Suzuki T, Suzuki Y, Hara Y, Shimamura T (1992). "The protective activity of tea catechins against experimental infection by Vibrio cholerae O1". Microbiology and Immunology. 36 (9): 999–1001. doi:10.1111/j.1348-0421.1992.tb02103.x. PMID 1461156. S2CID 34400234.
- ↑ Li S, Weidenfeld J, Morrisey EE (January 2004). "Transcriptional and DNA binding activity of the Foxp1/2/4 family is modulated by heterotypic and homotypic protein interactions". Molecular and Cellular Biology. 24 (2): 809–22. doi:10.1128/MCB.24.2.809-822.2004. PMC 343786. PMID 14701752.
- ↑ Smith RS, Kenny CJ, Ganesh V, Jang A, Borges-Monroy R, Partlow JN, et al. (September 2018). "Sodium Channel SCN3A (NaV1.3) Regulation of Human Cerebral Cortical Folding and Oral Motor Development". Neuron. 99 (5): 905–913.e7. doi:10.1016/j.neuron.2018.07.052. PMC 6226006. PMID 30146301.
- ↑ Vernes SC, Spiteri E, Nicod J, Groszer M, Taylor JM, Davies KE, et al. (December 2007). "High-throughput analysis of promoter occupancy reveals direct neural targets of FOXP2, a gene mutated in speech and language disorders". American Journal of Human Genetics. 81 (6): 1232–50. doi:10.1086/522238. PMC 2276341. PMID 17999362.
- ↑ Vernes SC, Newbury DF, Abrahams BS, Winchester L, Nicod J, Groszer M, et al. (November 2008). "A functional genetic link between distinct developmental language disorders". The New England Journal of Medicine. 359 (22): 2337–45. doi:10.1056/NEJMoa0802828. PMC 2756409. PMID 18987363.
- 1 2 Sia GM, Clem RL, Huganir RL (November 2013). "The human language-associated gene SRPX2 regulates synapse formation and vocalization in mice". Science. 342 (6161): 987–91. Bibcode:2013Sci...342..987S. doi:10.1126/science.1245079. PMC 3903157. PMID 24179158.
- ↑ Pennisi E (31 October 2013). "'Language Gene' Has a Partner". Science. Retrieved 30 October 2014.
- ↑ Smith K (11 November 2009). "Evolution of a single gene linked to language". Nature. doi:10.1038/news.2009.1079. ISSN 1744-7933.
- ↑ "Why can't chimps talk? It's more than just genes". Reuters. 11 November 2009. Retrieved 21 February 2019.
- ↑ Holy TE, Guo Z (December 2005). "Ultrasonic songs of male mice". PLOS Biology. 3 (12): e386. doi:10.1371/journal.pbio.0030386. PMC 1275525. PMID 16248680.
- ↑ French CA, Fisher SE (October 2014). "What can mice tell us about Foxp2 function?". Current Opinion in Neurobiology. 28: 72–9. doi:10.1016/j.conb.2014.07.003. hdl:11858/00-001M-0000-0019-F62D-4. PMID 25048596. S2CID 17848265.
- ↑ Wilbrecht L, Nottebohm F (2003). "Vocal learning in birds and humans". Mental Retardation and Developmental Disabilities Research Reviews. 9 (3): 135–48. doi:10.1002/mrdd.10073. PMID 12953292.
- 1 2 3 Teramitsu I, White SA (July 2006). "FoxP2 regulation during undirected singing in adult songbirds". The Journal of Neuroscience. 26 (28): 7390–4. doi:10.1523/JNEUROSCI.1662-06.2006. PMC 2683919. PMID 16837586.
- ↑ Heston JB, White SA (February 2015). "Behavior-linked FoxP2 regulation enables zebra finch vocal learning". The Journal of Neuroscience. 35 (7): 2885–94. doi:10.1523/JNEUROSCI.3715-14.2015. PMC 4331621. PMID 25698728.
- ↑ Jarvis ED, Scharff C, Grossman MR, Ramos JA, Nottebohm F (October 1998). "For whom the bird sings: context-dependent gene expression". Neuron. 21 (4): 775–88. doi:10.1016/s0896-6273(00)80594-2. PMID 9808464. S2CID 13893471.
- ↑ Bonkowsky JL, Chien CB (November 2005). "Molecular cloning and developmental expression of foxP2 in zebrafish". Developmental Dynamics. 234 (3): 740–6. doi:10.1002/dvdy.20504. PMID 16028276. S2CID 24771138.
- ↑ Hurst JA, Baraitser M, Auger E, Graham F, Norell S (April 1990). "An extended family with a dominantly inherited speech disorder". Developmental Medicine and Child Neurology. 32 (4): 352–5. doi:10.1111/j.1469-8749.1990.tb16948.x. PMID 2332125. S2CID 2654363.
- ↑ Gopnik M (September 1990). "Genetic basis of grammar defect". Nature. 347 (6288): 26. Bibcode:1990Natur.347...26G. doi:10.1038/347026a0. PMID 2395458. S2CID 4323390.
- ↑ Gopnik M (April 1990). "Feature-blind grammar and dysphagia". Nature. 344 (6268): 715. Bibcode:1990Natur.344..715G. doi:10.1038/344715a0. PMID 2330028. S2CID 4360334.
- ↑ Cowie F (1999). What's Within?: Nativism Reconsidered. New York, US: Oxford University Press. pp. 290–291. ISBN 978-0-1951-5978-3.
- ↑ Jenkins L (2000). Biolinguistics: Exploring the Biology of Language (Revised ed.). Cambridge, UK: Cambridge University Press. pp. 98–99. ISBN 978-0-5210-0391-9.
- ↑ Vargha-Khadem F, Watkins K, Alcock K, Fletcher P, Passingham R (January 1995). "Praxic and nonverbal cognitive deficits in a large family with a genetically transmitted speech and language disorder". Proceedings of the National Academy of Sciences of the United States of America. 92 (3): 930–3. Bibcode:1995PNAS...92..930V. doi:10.1073/pnas.92.3.930. PMC 42734. PMID 7846081.
- ↑ Fisher SE, Lai CS, Monaco AP (2003). "Deciphering the genetic basis of speech and language disorders". Annual Review of Neuroscience. 26: 57–80. doi:10.1146/annurev.neuro.26.041002.131144. hdl:11858/00-001M-0000-0012-CB7E-4. PMID 12524432. S2CID 1276712.
- ↑ "Genes that are essential for speech". The Brain from Top to Bottom. Retrieved 31 October 2014.
- ↑ Vernes SC, Nicod J, Elahi FM, Coventry JA, Kenny N, Coupe AM, et al. (November 2006). "Functional genetic analysis of mutations implicated in a human speech and language disorder". Human Molecular Genetics. 15 (21): 3154–67. doi:10.1093/hmg/ddl392. hdl:11858/00-001M-0000-0012-CB23-C. PMID 16984964.
- ↑ Feuk L, Kalervo A, Lipsanen-Nyman M, Skaug J, Nakabayashi K, Finucane B, et al. (November 2006). "Absence of a paternally inherited FOXP2 gene in developmental verbal dyspraxia". American Journal of Human Genetics. 79 (5): 965–72. doi:10.1086/508902. PMC 1698557. PMID 17033973.
External links
- Gene information at NCBI
- Gene information at Genetic Home Reference
- Language and Genetics Research at the Max Planck Institute for Psycholinguistics
- The FOXP2 story
- Revisiting FOXP2 and the origins of language
- FOXP2 and the Evolution of Language
- FactorBook FOXP2
PDB gallery | |
|---|---|
|